page 1/4
Quelques remarques
importantes
J'ai essayé d'illustrer mon propos par des schémas récapitulatifs ou informatifs. Si vous appréciez cette page, n"hésitez pas ŕ la signaler! Il est possible que j'édite dans quelques mois un CDrom intitulé "les origines" qui reprendra avec davantage de photos et schémas les sections astro et exobiologie de ce site.
Le problčme des conditions initiales
Nous, biologistes, ne tenons pas assez compte dans nos conceptions et descriptions de l'état prébiotique des conditions physiques du milieu: non seulement sa température, mais également la pression qui y rčgne, les champs magnétiques et électriques présents, les effets des rayonnements énergétiques, la composition chimique de l'atmosphčre primordiale... autant de domaines dans lesquels il faudra progresser. On a trop longtemps supposé que les conditions initiales était celles prévalant au sein des douillets rivages d'un océan tičde, surmonté d'une calme atmosphčre réductrice... Ces derničres années ont remis profondément en cause ce schéma: la vie s'est développée si vite que son milieu d'apparition devait ętre, selon nos critčres, apocalyptiques: sur des rivages irradiés d'UV, dans une atmosphčre étouffante de dioxyde de carbone, soumise aux émissions gazeuses de fréquents épanchements de lave, aux chutes de météores et de noyaux cométaires de tout diamčtre, ŕ un continuel apport de matičre organique synthétisée dans l'espace et ŕ des modifications trčs rapides des conditions physiques, en particulier en ce qui concerne la température.
Si vous avez oublié d'écouter en cours de bio: une remise ŕ niveau expresse, histoire de savoir de quoi on parle...
|
Il est des espaces sis entre les étoiles Etrangers ŕ nos yeux, solitudes d'atomes Ou la lumičre joue en ses champs magnétiques Imprimant dans la pierre les briques de la vie Et nous voyons poussičres, y compris dans nos livres et notre espoir toujours, en vaine éternité devant nous dévoile, au dessus de nos tętes l'histoire de l'origine... RR |
 |
Sommaire
Des atomes aux molécules organiques
Les aventures de la polymérisation - la crise de l'énergie
Le ribomonde (Ou l'ARN et les peptides prennent le relais)
5 - Conclusion: la vie est une réaction en chaîne ŕ l'échelle galactique
Le top départ : oů l'on apprend comment répondre ŕ Mr Fermi
| élément |
|
|
|
|
|
|
|
|
|
|
|
|
| % masse |
|
|
10 |
|
|
|
|
|
0,08 |
|
|
0,009 |
| élément |
|
|
|
|
|
|
|
|
|
|
|
|
| % masse | 0,005 | 0,002 | 0,002 |
|
|
|
|
0,00005 | 0,00002 | 0,000003 | 0,0000005 |
|
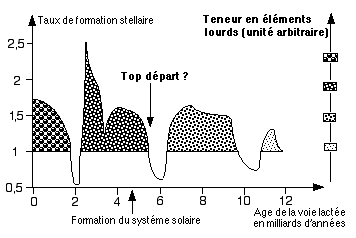
Pour notre galaxie, la voie lactée, les nuages moléculaires et la plupart des étoiles de grande masse se situent dans le plan de la galaxie. On suppose que des perturbations d'origine gravitationelles, impliquant peut ętre les nuages de Magellan, deux galaxies satellites de la notre, ont été ŕ l'origine de 3 ou 4 périodes de formation stellaires (voir schéma ci contre) impliquant 3 époques d'enrichissement du milieu interstellaire en éléments lourds (autres que H et He). On peut considérer que ces éléments ont été disponible il y a 5 ŕ 6 milliards d'années environ. C'est ŕ cette époque que je situe le "top départ" stellaire pour l'apparition du "phénomčne vivant". Cela
nous fournit une date
butoir, bien que trčs
imprécise, pour
l'apparition de la vie dans notre galaxie.
|
3 vagues de formations stellaire ont été identifiées dans notre galaxie, chacune enrichissant le milieu interstellaire en éléments lourd (schéma personnel) |
A partir du moment ou nous trouvons dans l'environnement carbone et oxygčne en abondance, la chimie du carbone peut se mettre en route. Elle nécessite un support: la formation d'astéroďdes, de poussičres et de plančtes telluriques est une condition supplémentaire pour notre "top départ" que l'on peut donc estimer autour de 5 milliards d'années. Cette idée que la disponibilité des éléments lourds est une condition clef de l'évolution des systémes stellaires vers la formation de plančtes, puis de formes de vies, est corroborée par plusieurs observations:
Si l'on tient compte des conditions "locales" ŕ l'échelle galactique (onde de chocs en provenance de supernovae proches, vagues de formation stellaire, enrichissement du milieu interstellaire....), nous voyons que pour une large région de la galaxie les conditions "stellaires" liées ŕ l'apparition de la vie sont grosso modo synchronisées. On peut donc en déduire que la plupart des civilisations extraterrestres potentielles voisines sont peu ou prou ŕ un niveau similaire. Voila peut ętre pourquoi, Mr Fermi, ils ne sont pas (encore) lŕ....
Toutefois, certaines observations, comme celle du Quasar APM 8279+5255, distant de 13.5 milliards d'AL et donc tout aussi vieux, montrent que ces derniers contiennent déjŕ des quantités importantes de fer (3 fois supérieures ŕ celles du systéme solaire pour cet exemple - Hasinger G, 2002). La vitesse de synthčse des éléments lourds pourrait donc avoir été trčs différente dans des zones diverses de l'univers.
Pour ce qui nous concerne, le départ dans notre région de la voie lactée semble situé il y a 4,5 milliards d'années...
Des atomes aux molécules organiques
Nous
sommes dans la région de la galaxie va naître le
soleil, il y a 5 milliards d'années.  Le
systčme solaire ressemble alors
ŕ
une espčce d'oeuf au plat: un protosoleil central et un
disque d'accrétion, peut ętre fragmenté
en
anneaux, qui tournent autour de lui. Cette disposition est assez
fréquente, et nombreuses sont les étoiles qui
sont
entourées de plančtes ou de disques
d'accrétions
(b pictoris, 68
Ophiuchi, BD 31643).
Le
systčme solaire ressemble alors
ŕ
une espčce d'oeuf au plat: un protosoleil central et un
disque d'accrétion, peut ętre fragmenté
en
anneaux, qui tournent autour de lui. Cette disposition est assez
fréquente, et nombreuses sont les étoiles qui
sont
entourées de plančtes ou de disques
d'accrétions
(b pictoris, 68
Ophiuchi, BD 31643).
Les molécules présente alors dans ce nuage sont plus complexes que l'on pourrait le croire. Certes, au gré des chocs, le carbone se combine avec les autres atomes présents (surtout de l'hydrogčne), mais surtout la présence de poussičres fournit un substrat sur lequel se produisent des réactions chimiques qui utilisent l'énergie du rayonnement stellaire. Ces poussičres (de 0,1mm ou moins), en rapprochant les atomes et les molécules avant leur interaction, jouent le męme rôle que les catalyseurs minéraux (mousse de platine par exemple) utilisés en chimie. Les molécules formées résultent principalement de combinaison avec de l'hydrogéne. Ces poussičres forment des structures floconneuses comportant de la glace amorphe de haute densité, se comportant comme un fluide visqueux, des silicates et des molécules ŕ base de carbone (hydrocarbures principalement). Ces particules vont s'aggréger pour former deux types de corps solides: les plančtes telluriques prčs du soleil et, plus loin, les noyaux des futures comčtes. Entre les deux, les éléments légers chassés par la pression de radiation du jeune soleil vont se condenser sur des "noyaux" telluriques pour former les plančtes joviennes, nanties de nombreux satellites aux dimensions respectables.
Ci dessus:
occultation
de l'étoile binaire BD31643, qui permet
de mettre en
évidence
son disque protoplanétaire
(photo
Kalas P & Jewitt D)
Quelles sont les molécules qui peuvent ętre synthétisées dans les conditions "interstellaires" et que nous retrouverons au début de la formation du systčme solaire? Tout dépend des nuages moléculaires ŕ l'origine du systčme solaire. Il en existe deux types principaux dans les bras spiraux de la voie lactée:
|
On
peut
identifier les molécules présente dans ces nuages
par
leur spectre, en observant actuellement les grands nuages
moléculaires qui parsčment le disque de notre
galaxie (et
des autres!). Bien qu'a l'époque de la formation du soleil
la
teneur en éléments lourds devait ętre
un peu moins
élevée, cela nous donne une indication sur les
apports de
molécules extraterrestres qui ont pu ętre
réalisées au début de l'histoire de
notre
plančte.
On
y trouve plus de 120
molécules
différentes, dont
surtout de l'eau,
du
méthane, de l'ammoniac, du monoxyde
de carbone, des
radicaux libres (OH, H3+) intervenant dans
les synthčses des autres
molécules, mais également des
molécules plus
complexes.
|
 Structure d'un grain de poussičre interstellaire... |
|
Des hydrocarbures (HC3N, CH3CN, CH3CHO...), des alcools (méthanol, éthanol) et des dérivés du cyanure se forment aussi facilement: les glaces qui recouvrent les silicates, composées d'eau, de méthanol et d'hydrocarbures, jouent le rôle de pičge moléculaire qui empęche la dispersion des molécules qui peuvent se recombiner sous l'action du rayonnement UV stellaire. Un nuage de gaz situé prčs du centre galactique a męme révélé contenir du glycolaldéhyde (C2H4O2) qui peut en se combinant conduire ŕ du glucose ou du ribose (P. Jewell. 08/2000) . Ce sucre a également été détecté dans des météorites, accompagné de composés comme le glycérol, présents dans les membranes de cellules. (Cooper & al., 2001). Le
processus de formation des
molécules peut parfois nécessiter plus de 1500
étapes intermédiaires, mais met souvent en jeu
l'ion H3+
qui se combine avec les autres éléments
présents
dans le milieu interstellaire.
|
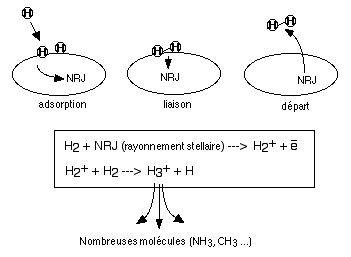 Formation d'hydrogčne moléculaire et d'ion intermédiaires dans les réactions de synthčse interstellaire |
|
On
peut s'étonner que
dans des zones ŕ basse température (espace), des
réactions chimiques puissent se produirent facilement. En
fait,
dans ces conditions, la cinétique des réactions
n'obéit plus ŕ la loi d'Arrhénius: la
basse
température ralentit l'agitation moléculaire et
atomique,
ce qui permet aux forces électrostatiques, importantes
puisque
les molécules interstellaires sont souvent
chargées
(radicaux libres) d'agir pour rapprocher les
éléments
devant réagir (cf Pour
la science 266, 12/1999, p 14).
Les études expérimentales de la formation de molécules dans le milieu interstellaire (Mayo Greenderg, 1984, Allamandola, 1987, Berstein, 1999) montrent qu'a partir des molécules présentent ŕ l'état gazeux entre les particules solides il se forme des couches successives de matériaux contenant de nombreuses molécules carbonées: il y a incorporation du carbone dans une "matrice contenant de l'eau sous la forme de glace amorphe (non cristallisée).
|
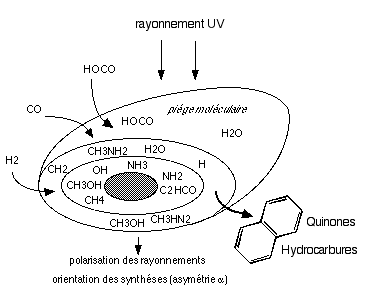 Les grains de poussičre jouent dans l'espace le rôle d'intermédiaires de synthčse, de catalyseurs... et sur Terre ? |
|
Origine
de la poussičre
interstellaire La
plupart des particules de
poussičre interstellaire prennent naissance ŕ
partir des
particules de silicates qui se condensent dans l'atmosphére
des
étoiles supergéantes de classe M. La pression de
radiation causés par le rayonnement stellaire propulse dans
l'espace ces grains, leur température n'étant
plus alors
que de l'ordre de la dizaine de Kelvin (- 263 °C). Initialement
amorphes, ils adoptent alors une configuration partiellement ou
totalement cristalline (Molster
& al., 1999). C'est
ŕ partir de ces particules que vont se condenser les grains
interstellaires. Bien que ces processus se déroulent dans un
environnement trčs peu dense, les grains grossissent en
incorporant puis en modifiant les éléments qu'ils
rencontrent.
|
Les molécules du systčme solaire primitif
Gardons présent ŕ l'esprit que les silicates composant les grains de poussičre contiennent une proportion importante de sulfures métalliques, d'oxydes et d'argiles. De part leur structure floconneuse, ces composés offrent une énorme surface sur laquelle des synthčses peuvent se développer. Ils ont une grande importance pour la suite des événements.
Alors
que se forment
plančtes,
comčtes et astéroďdes, la
nébuleuse
protosolaire contient donc déjŕ de nombreuses
molécules
organiques: eau, alcools, acides aminés... ainsi que des
catalyseurs potentiels. Température, densité et
pression diminuent au fur et ŕ mesure que l'on
s'éloigne
du soleil.
Nous sommes il y a 5 milliards d'années. Pour l'essentiel, les plančtes se sont formées et sont en train de se refroidir. Les plančtes telluriques, chauffées sous le triple effet de leur contraction gravitationelle, de la désintégration de leurs éléments radioactifs et des impacts de météores, voient leurs éléments les plus lourds se rassembler pour former rapidement (quelques dizaines de millions d'années) un noyau, surmonté d'un manteau fluide et d'une croűte en cour de solidification, entourée d'une atmosphčre principalement composée de dioxyde de carbone et d'un peu d'azote, atmosphčre sans doute trčs épaisse (Allégre, 1994) et plus riche en hydrogčne que l'on ne l'aurait cru.
Les analyses menées en avril 2005 par la sonde Cassini sur Titan montrent que la haute atmosphčre constitue également une source non négligeable de molécules organiques : sous l'influence du roiayonnement solaire, les molécules de N2, CO2, CH4 et H2 se recombinant. Sur Titan, actuellement, ces réactions aboutissent ŕ la formation de molécules comportant 7 atomes de C (soit plus que les bases puriques et pyrimidiques !) qui "sédimentent" ensuite vers le sol.
Aspect d'un grain de poussičre interstellaire en microscopie électronique a balayage: la structure floconneuse apparaît nettement. Au niveau des dimensions et de la forme, cela correspond aux ribosomes actuels. Photo JP Bradley
| Jusqu'ŕ
3,9 milliards
d'années,
les collisions dans le systčme solaire étaient
extręmement fréquentes
(Bendjoya, 1998).
L'une d'elle, il y a 4,5 milliards d'année, avec un corps de la taille de Mars, a été si violente qu'une grande partie de l'enveloppe externe de la Terre a été arrachée, s'est satellisée et a formé la Lune. A cette occasion, notre plančte a perdu sa premičre atmosphčre et ses océans éventuels, se trouvant recouverte d'un océan de Magma d'un millier de km de profondeur (Righter & al., 1997). Ces impacts géant vont avoir des conséquences sur l'atmosphčre qui va se former: le coeur de fer de notre plančte va se former trčs rapidement, en moins de 30 millions d'années (Yin et al. 2002), il ne percolera pas ŕ partir du manteau, ce qui influera sur la composition chimique de ce dernier et les gaz qui vont d'en échapper, lesquels ne seront pas aussi réducteurs que le croyait les premiers chercheurs, comme Urey, ŕ s'ętre interessé au probléme dans les années 60. L'incessant
bombardement qu'a subi
la Terre semble coincider avec le développement de la vie:
loin de bénéficier de calmes océans,
il semble
bien que la vie ait commencé dans des conditions chaotiques!
Entre 4 et 3,85 milliards d'année, la Terre a subie 20
fois plus d'impacts que la lune (Zahnle
& al., 1997).
Malgré l'atmosphčre,
10 ŕ 20 de ces impacts ont du creuser des bassins
de plus de 2000 km de diamčtre et ont
dégagés
assez d'énergie pour vaporiser toute l'eau de la
plančte.
Il est toutefois possible, comme le signale G. Ryder (2003) que la
chronologie des impacts majeurs, fondés sur l'analyse et la
datation des structures lunaires, soit en réalité
trčs floue et que ces impacts cataclysmiques,
stérilisateurs, aient eu lieu il y a plus de 4 milliards
d'années, laissant ŕ la vie un milieu
agité
certes, mais moins délčtčre que celui
couramment
envisagé (pendant les derniers 600 millions
d'années,
1500 astéroďdes de plus d'un km de
diamčtre ont
frappé la Terre, dont 200 sur les continents: certain ont
provoqué des extinctions, mais aucun n'a détruit
toutes
les formes de vies qui ont évoluées sans cet
intervalle...). Toutefois, il semble bien que l'on doive conclure que
la vie apparaît sur Terre dčs que cessent les
bombardements les plus
importants (ceux impliquant des corps de plusieurs centaines de km de
diamčtre). |
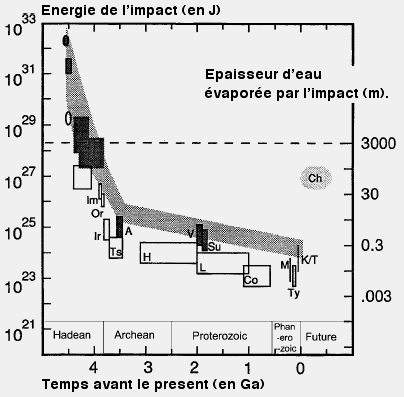
Energie libérée par les impacts météoritiques subis par notre plančte. Les carrés vides se réfčrent ŕ des données lunaires (tycho, Copernic, Langrenus, Hausen, Tsiolkovsky, Iridum, mare orientale et mare Imbrium) alors que les pleins documentent des impacts terrestres avérés (Chicxulub, Manicugan, Sudbury, Vredevort et l'épaisseur des différentes sphériules datant de l'Archéin. La premičre donnée correspond ŕ la formation de la Lune. La bande grisée décrit bien la décroissance de l'énergie destructrice des impacts. La ligne pointillée signale la limite en deça de laquelle un impact ne vaporise plus toute l'eau des premiers océans. Adapté de Sleep & al., 1989 (Sleep NH, Zahnle KJ, Kasting JF, Morowitz HJ. 1989. Nature 342:139–42) |
Par la
suite, alors que
l'activité volcanique libérait
de nombreux gaz, dont de la vapeur d'eau, les comčtes
enrichissaient
également la Terre en eau alors que les
astéroďdes
et surtout les micrométéorites (Maurette, 1998) se chargeaient
d'amener des
composés
non seulement carbonés mais aussi azotés (Marty &
al., 2001). Il en
était de męme
pour toutes les plančtes telluriques (exceptée
Mercure,
trop prés du soleil). En effet, et contrairement
ŕ la
théorie encore majoritairement acceptée par les
géologues (et émise pour la premičre
fois en
1894!), les matériaux (fer et silicates) qui ont
formé
notre plančte ne pouvaient pas
contenir originellement assez de
matériaux volatils pour constituer une atmosphčre,
car
ils ont été porté ŕ une
température
de l'ordre de 1000 K (Delsemme, 1996; d'aprčs Larimer et
Anders).
|
Des données récentes amčnent ŕ reconsidérer la dynamique du développement du vivant ŕ l'échelle galactique. Ces dix derničres années, nous avons appris que:
Il en résulte que si l'on rapproche le taux de développement des bactéries, l'époque de l'apparition de la vie et la fréquence des impacts météoritique, ils est trčs probable que plusieurs météores comportant des formes de vies primitives aient pu ętre expédiés au hasard au delŕ du systčme solaire ou ŕ l'intérieur de celui ci. Ces formes de vies se révélant parfaitement capable de résister ŕ un long voyage et ŕ une rentrée atmosphérique, on doit en déduire que les plančtes du systčme solaire (ainsi que peut ętre, les étoiles proches), ne sont pas biologiquement isolées. Les conséquences de ce phénomčne de dissémination sont importantes:
|
Les aventures de la polymérisation
L'irritant problčme de la chiralité du vivant
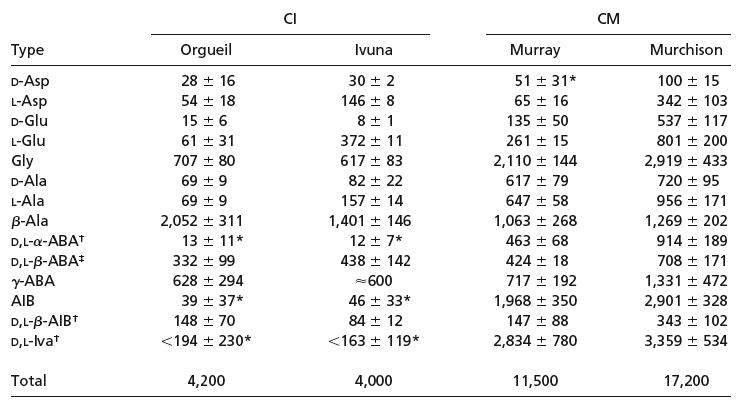
Cerrtains auteurs sont malgré tout ŕ la recherche d'un milieu de synthčse assymétrique terrestre: Ainsi, il a été proposé (Goodman & Gershwin, 2006) que la rotation (assurément asymétrique !) de la Terre provoque, ŕ l'aube et crépuscule, en opposition de phase avec les profils de température (inconnues...) des océans, un tri isomérique parmi les acides aminés: les isoméres D des AA aurait été détruits au crépuscule dans une mer plus chaude qu’ŕ l’aube, produisant ainsi un excés quotidien d’isoméres L protégé du rayonnement par la nuit (en négligeant l'influence de la Lune...) et par sa diffusion dans les zones océaniques plus froides et plus obscures. les premičres formes de vies marines auraient incorporé cet excés isomérique fixé depuis dans le vivant.
Pour expliquer l'homochiralité D des acides nucléiques également faire appel ŕ des précurseurs de l'ARN, les pyranosyl-ARN dont la plus grande résistance et la meilleure sélectivité pourrait expliquer cette sélection isomérique (Bolli et al., 1997). On ne peut cependant exclure un effet provenant de certains catalyseurs minéraux terrestres, mais cela semble ŕ ce jour bien improbable. Aucune explication convainquante de cette homochiralité n'a encore été avançée.
Certaines études avait fait apparaitre que les acides nucléiques ne pouvaient conserver leur propriétés de réplication en présence d'une population racémique d'acides nucléiques. Toutefois, l'importance de cette inhibition croisée énantiomérique a été grandement exagérée : elle ne s'exerce que vis ŕ vis de monomčres, alors que l'addition d'oligomčres permet de contourner cette difficulté (Borcher & al., 2004). Dans les conditions prébiotiques, l'existence d'un milieu comprenant un mélange de monomčres et d'oligomčres est probable, ce qui rend peu plausible un "blocage" en milieu racémique.
Une minirevue (en anglais) parue en 2004 dans Experimental biology and medicine
Les
différents lieux de
l'apparition de
la vie
A la surface des plančtes telluriques...
Il y a 4 milliards d'années, la croűte terrestre est ŕ peine refroidie. Notre plančte s'est formée, d'aprčs les donnée radiométriques et l'étude des météorites, il y a 4,56 ± 0,01 Ga, par un processus d'accrétion qui a duré environ 100 millions d'années (Canup & al., 2000).
Les éruptions volcaniques sont fréquentes, la température des océans voisine de 80°C. Une épaisse atmosphčre (80% d'eau, 19% de CO2 et 1% d'azote) enveloppe la Terre, mais aussi probablement Vénus et Mars, ou plusieurs grandes étendues d'eau atteignent plus de 10m de profondeur.
|
L'absence
d'oxygčne, donc d'ozone, expose la surface des
plančtes
au rayonnement UV solaire (absorbé par 10 m d'eau environ),
bien
que notre étoile soit alors au minimum de sa
luminosité (30% inférieure ŕ
l'actuelle).
Cette
situation n'est pas sans poser problčme: notre
plančte
aurait du ętre recouverte de glace, ce qui implique
l'existence
d'un trčs fort effet de serre pour maintenir une
température élevée (ŕ moins
que la vie ne
soit apparue sur une terre ressemblant ŕ une boule de neige,
ce
qui est effectivement arrivé bien plus tard dans l'histoire
de
notre plančte - Bodiselitsch & al., 2005),
attestée
par la présence d'eau liquide qui a laissé sa
marque dans
les roches de cette lointaine époque.
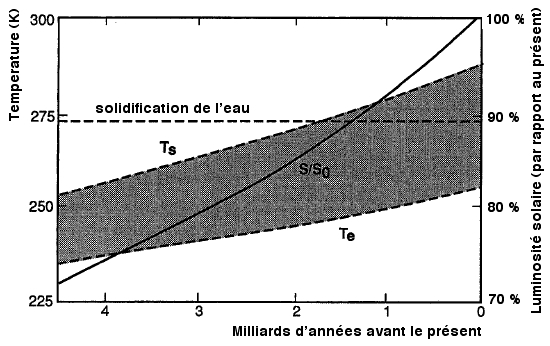
Le graphique ci contre,
(adapté de
Kasting & al.,1988 d'aprčs Gough, 1981) illustre les
modifications de la luminosité solaire au cours de
l'histoire de
la Terre.
La
courbe
représente l'augmentation de la luminosité
solaire en
fonction du temps, alors que la zone grisée est comprise
entre
un minima correspondant ŕ la température
radiative de la
Terre et un maximum correspondant ŕ la
température
moyenne de la plančte (calculs effectués sur un
modčle ou CO2 et H2O ont des valeurs correspondante
ŕ 300
ppmv CO2).
|
 |
La Lune, 3 fois plus proche de la Terre et plus inclinée sur l'ecliptique qu'actuellement, y provoque des marées de forte amplitude (10 fois le niveau actuel), et ce d'autant plus fréquemment que la rotation de notre plančte ne dure que 12 h seulement! Les pluies sont trčs acidifiées par le CO2 atmosphérique et attaquent les silicates, amenant ainsi de nombreux ions en solution dans une eau trčs chaude...
|
ARGUMENTS
H2O liquide sur Terre avant 3 Gy: - Sédiments aquatiques entre 3.8 et 3.9 Ga (A. P. Nutman, J. H. Allaart, D. Bridgewater, E. Dimroth, M. Rosing, Precambrian Res. 25, 365, 1984) - Stromatolithes dans des zones de maréeil y a 3.5 Ga - laves en coussin formées par des magmas submergés agées de plus de 3 Gy. Depots détritiques de grains d'uraninite entre 3.0 et 2.3 Ga - turbidites et traces sous marines entre 3.5 et 3.0 Ga (K. A. Eriksson, Tectonophysics 81, 179, 1982). |
ARGUMENTS
Terre a climat chaud avant 2,5 Gy:
- Absence d'indices de glaciation avant 2.7 Ga (J. F. Kasting, Palaeogeogr. Palaeoclimat. Palaeoecol. 75, 83, 1989)- Possibilité que les tillites les plus recents ait été formées par des impacts plutot que par des conditions glaciaires, ce qui implique l'existence d'un climat chaud voici plus de 2 Ga (V. R. Oberbeck, J. R. Marshall, H. R. Aggarwal, J. Geol. 101, 1,1993) - Thermophilie des archaebacteria [C. Woese, Microbiol. Rev. 51, 221,1987) mais qui peut corespondre ŕ un caractére sélectionné dans une population qui a survécu ŕ une période de hate température résultant d'un des derniers impacts majeurs il y a 3,8 Gy (ce qui implique une origine trčs précoce de la vie) - M. Gogarten- Boekels, E. Hilario, J. P. Gogarten, Origins Life 25, 251, 1995) - L'étude des isotopes du soufre indique une température océanique comprise entre 30 et 50°C entre 2.6 et 3.5 Ga (H. Ohmotu, R. P. Felder, Nature 328, 244, 1987) - Les isotopes de l'oxygčne indiquent une température moyenne de surface de plusieurs dizaines de °C entre 2.5 et 3.5 Ga (L. P. Knauth, S. Epstein, Geochim. Cosmochim. Acta 40, 1095, 1976). - D'autres données comparant les taux d'isotope 18 de l'oxygčne dans des cherts et des phosphates de la męme époque indiquent des températures de l'ordre de 100°C voici 3.8 Ga [J. Karhu, S. Epstein, Geochim. Cosmochim. Acta. 50,1745, 1986). |
A partir des molécules organiques synthétisées in situ et celles d'origine extraterrestre, les premiers polyméres (peptides et acides nucléiques) vont se former. Cette polymérisation a lieu simultanément dans plusieurs milieux, ŕ des vitesse et avec des rendements différents:
Résumons nous: on se retrouve avec un monde chaud, nanti d'eau, d'acides aminés ŕ la pelle, d'adénine, mais ou l'ARN semble bien difficile ŕ faire apparaître...
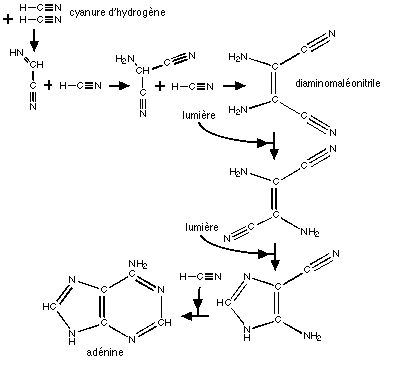
Un précurseur innatendu: l'Urée.
Le Dr N.Psychogios, pharmacien chimiste, a attiré mon attention sur un mode possible de formation des précurseurs des acides nucléiques utilisant l'urée (ce n'est pas la premičre fois: dés 1995, Robertson et Miller - oui, celui de l'expérience- ont proposé une voie de synthčse de cytosine et d'Uracyle ŕ partir d'Urée et de cyanoacétaldéhyde; voir ici pour plus de détails) . Il propose le scénario suivant (que j'illustrerai dčs que je le pourrai):
1 - De l'urée se forme ŕ partir d'ammoniac et du CO2 de l'atmosphče prébiotique. De l'ammoniac devait ętre présent dans les océans, et comme le précise N. Psychogios: " NH3 et CO2 ont du logiquement réagir en tant qu'acides et bases "dominants", précipiter sur les rives de l'océan primitif et se déshydrater pour donner de l'urée (solide ou liquide) qui elle męme était alors immédiatement disponible pour réagir avec l'atmosphčre. Le milieu local était forcément basique car l'urée se dimerise en milieu anhydre et chaud en liberant de l'ammoniac, lui męme sobluble dans l'urée, contrairement au CO2. ". L'urée se forme dans les expériences " ŕ la Miller", mais avec un rendement trčs faible (0,03 %)
Les conditions de la synthčse ici décrite sont une forte pression et une température élevée: 140 bar et 150 ŕ 180°C, mais elles ont pu ętre obtenues ŕ partir d'impacts, ou dans l'atmosphčre lors de l'ablation de météorites de faible masse. cette synthčse fait intervenir un intermédiaire, le carbamate d'ammonium, dont la déshydratation forme l'urée. Cette molécule est peut ętre une des cles des synthčses prébiotiques.
2- L'urée formée réagit ŕ son tour avec deux molécules dont on peut supposer qu'elle ait été présentes dans l'atmosphčre prébiotique: le cyanoacétylčne et le formylacétylčne (tous deux identifiés dans l'atmosphčre actuelle de Titan). La réaction qui se produit est une condensation, qui aboutit aux résultats suivants:
cyanoacétylčne + urée --> thymine
formylacétylčne+ urée --> uridine
Comme le remarque notre chimiste avec raison: " Les produits de condensation de l'urée sur le cyanoacetylene et le formylacetylene ne sont par contre pas ou peu solubles dans l'eau ce qui provoque immanquablement une concentration de ses bases en bordure d'ocean. " De plus, ces réactions ne font pas intervenir l'eau. Elles peuvent donc se produire "ŕ sec", oů dans des milieux pauvres en eau (soupe d'hydrocarbures, urée liquide... ?)
Cette voie conduirait ŕ une accumulation de bases dans un milieu "sec", donc plus favorable ŕ une polymérisation ultérieure que les scénarios faisant intervenir des réactions dans un milieu riche en eau.
The problčme: polymérisation des acides nucléiques...
Pour enchaîner les différents monomčres, il faut réaliser des réactions de polymérisation. Pour cela, il faut lier deux molécules en leur enlevant une molécule d'eau. Malheureusement, en milieu aqueux, c'est la dépolymérisation (hydrolyse) qui est favorisée (empęchant l'allongement des polymčres protéiques ou nucléiques). Comme le déclare Shapiro (1998): "L'eau n'avait de cesse d'empęcher l'assemblage des grosses biomolécules et de détruire celles qui étaient parvenues ŕ s'assembler". On doit donc trouver un milieu qui favorise la polymérisation. Pour cela, il faut concentrer les réactifs ou enlever l'eau, et surtout coupler la réaction ŕ une source d'énergie (par exemple une autre réaction qui libérerait de l'énergie). En effet, l'allongement d'un polymčre nécessite de l'énergie alors que l'hydrolyse produit de l'énergie, ce qui lui permet de se produire spontanément. Comment résoudre ce problčme, parfois pudiquement évacué par les meilleurs "bio-men "?
Je propose une synthčse des prč-ARN catalysés par et sur les poussičres interstellaires porteuses d'acides aminés. Des acides nucléiques pourraient aussi ętre synthétisés au niveau des grains de poussičre interplanétaire, polymérisés sous l'effet des rayonnements et adsorbés ŕ leur surface. L'expérience de cette synthčse lente, ŕ froid et ŕ sec, reste ŕ faire.
Il est
cependant possible
d'obtenir une synthčse de nucléoside puriques
(ribose
+ base purique mais sans phosphate - Orgel,
1994). On obtient ainsi
une grande variété de molécules ("ARN-like") de
pré-ARN
puriques
(comprenant A et G).
En effet, il est difficile d'obtenir la
liaison chimique entre le ribose et les pyrimidines (C et U). Plusieurs
autres indices venant de l'observation du métabolisme actuel
de
nos cellules plaident, qui plus
est, en faveur de la formation de
nucléotides primordiaux essentiellement puriques.
Il
serait possible que les premiers
nucléotides aient simplement
été des chaines de purines (xanthines ?)
liées
ŕ des
ribose de forme furanose. Ces molécules, ne pouvant
s'apparier,
auraient été au départ "simple brins",
liées et stabilisées par exemple par des
éléments minéraux. Un des point
d'achoppement
principaux consiste ŕ réaliser la liaison entre
les
nucléotides au moyen d'un groupe phosphate. C. de Duve
propose
une polymérisation utilisant l'énergie de la
liaison
thio-ester et des phosphates fournit par une eau suffisemment acide (ce
qui était le cas avec une atmosphčre de 80% CO2)
pour
dissoudre le phosphate de Calcium des roches.
Les courtes chaines de pyranosyl ARN sont capable en solution de se lier, en effectuant une sélection des diverses formes optiques, et de former ainsi des oligoméres se replient et se liant comme un ARN "ŕ ribose". Plus simplement encore, les molécules de la famille des oligonucleotides (L)-a-threofuranosyl (TNAs), dérivés d'un sucre ŕ 4 carbone seulement, sont capables de s'apparier et de se lier ŕ la fois avec l'ARN et l'ADN (Schoning et al., 2000). Ces molécules représentent donc de bon candidat au titre d'ancétre des nucléotides! Autres candidats moléculaires au titre d'ancętre de l'ARN, les PNA (peptidic nucleic acid) oů acides peptidiques nucléiques (Nielsen, 2004) dans lesquels les riboses de l'ADN sont templacés par des acides aminés. Ces molécules situées ŕ mi chemin entre protéines et acides nucléiques, sont étudiées pour les éventuels effets thérapeutiques mais sont également capable de s'apparier en doubles hélices trčs stables, et présentent un comportement reproduisant bien celui de l'ADN.
En effet, la capacité de former des appariement antiparalléles (paires de Watson-Crick) est assez répandue chez les nucléotides. Cela permet d'étudier leurs propriétés lors de synthčses in vitro, et de les comparer ŕ celles des ARN actuels. D'autres édifices moléculaires peuvent présenter des capacités de reproduction, d'évolution et d'exploitation d'un milieu (Rebek, 1994). Ainsi, une molécule comme l'A.R.N.I. (adénine-ribose-naphtalčne-imide) est capable de se reproduire. Sous l'influence de rayonnement UV, une population de molécules légérement différentes se constitue: on retrouve une quasi espčce. Une autre molécule, la di-aminotriazine xanthčne-thymine (DIXT) présente également cette capacité d'évolution. On peut notre que l'adénine entre fréquemment dans leur composition, ce qui est interessant car un mécanisme de synthčse pré-biotique de cette molécule est connu. Les autres molécules utilisées peuvent ętre synthétisées sur les grains de poussičre interstellaire...
|
Une
hypothčse iconoclaste: le tout protéine
Et
si nous faisions fausse
route ? Depuis la découverte de T. Cech, tout le monde
considčre que l'ARN est la premičre
molécule
organique originelle. Et si nous avions eu au début une
information protéique,
bien plus simple, se transmettant
simplement par contact avec
des oligonucléotides ?
Cette idée m'est venue en effectuant des recherches sur le
fameux prion, cette protéine extręmement
résistante
(hélas!) capable de transmettre son "information" sans
matériel nucléotidique.... La premičre
molécule autoréplicative n'aurait elle pas pu
ętre
un "paléoprion", transmettant uniquement une
forme donc
découlerait, plus tard, une fonction... Je pose la question.
Historiquement,
la piste des protéines a
précédé celle des
acides nucléiques. Elle a été
particuličrement explorée par l'équipe
de S. Fox,
directeur de l'institut de génétique
moléculaire
et cellulaire de Miami, disparu en 1998. Ce dernier a obtenu par
chauffage d'un mélange d'acides aminés divers des
polymčres voisins des protéines, les
protéinoďdes, qui présentent la
propriété de pouvoir former des
sphéroďdes
microscopiques capables de se diviser, mimant en cela des processus
biologiques et fournissant la matičre premičre
ŕ
l'édification de "capsules" protéiniques
protégeant les premiers organismes...
Dans
les années 80, l'équipe de Stuart Kauffman a
envisagé la possibilité d'une vie primitive
essentiellement constituée d'un ensemble de peptides
douées d'activités autocatalytiques (Kauffman;
Farmer
& al., 1986). Le gčne serait alors apparu
aprčs les
protéines, dont les acides aminés sont obtenus
par les
synthčses prébiotiques. L'approche originale de
cette
équipe consiste ŕ considérer qu'il n'y
a pas eu
une molécule comparable d'emblée ŕ
l'ADN mais
émergence d'un comportement d'ensemble, concernant une vaste
population moléculaire peptidiques (comportant entre 5 et 27
AA), reproduisant celui dévolu actuellement aux acides
nucléiques. Ainsi, la reproduction de ce "pool
autocatalytique" est accomplie car chaque
espéce
moléculaire catalyse un peu la formation d'une autre,
laquelle
influe ŕ son tour sur une troisičme... ect. Ce réseau,
une fois
modélisé informatiquement, montre
alors que la reproduction et l'évolution
moléculaire sont
possibles dans ce cadre. Le transfert de l'information ŕ un
acide nucléique serait intervenu plus tardivement,
permettant de
multiplier le nombre de molécules peptidiques possibles. Ce
schéma nous raméne aux idées de J.
Keosian et du physicien F. Dyson selon laquelle "les premiers
fossiles seraient ceux d'organismes qui fonctionnaient avec un
systéme génétique fondé sur
les
protéines, fonction transférée
secondairement aux
ARN puis ŕ l'ADN".
|
L'indispensable
mécanisme
de couplage énergétique
Un
apport d'énergie est
indispensable ŕ la formation des liaisons chimiques liant
les
monomčres entre eux. Cette énergie est fournie
par des
liaisons chimiques doubles ou triples, dans des molécules
dites
"riches en énergie". Ces molécules (des
anhydrides)
présentent l'avantage de se former spontanément
en milieu
chaud et anhydre (Baltscheffsky). Certains composés comme
le carbodiimide (-N=C=N-) fournissent assez d'énergie pour
une polymérisation. D'autres agents de couplage sont le
cyanogčne, le cyanamide, les
cyanoacétylčne
et le diaminomaléodinitrile (leur énergie est
stockée
dans une triple liaison CC ou CN).
Les
pyrophosphates, ou phosphates
minéraux, ont pu également fournir au monde
prébiotique une source d'énergie
appréciable (et
qui expliquerait pourquoi tous les organismes utilisent toujours dans
leur métabolisme l'énergie contenue dans l'ATP
ou,
parfois, le GTP). En effet, des bactéries
photosynthétiques stockent toujours leur énergie,
grace
ŕ une "PPase", dans des phosphates minéraux alors
que la
photosynthčse des eucaryotes met en jeu une ATPase et de
l'ATP
(cette photosynthčse bactérienne semble
adaptée
ŕ une forte intensité lumineuse et ŕ
une
disponibilité moindre des ions H+). De plus, ces
pyrophosphates
sont produits, par exemple, pendant les éruptions
volcaniques:
Il semble que, trčs rapidement, les premiers métabolismes (intégrés dans les ętres vivants oů les précédant) se soient caractérisés par des mécanismes permettant d'accumuler de façon inégale des ions, ceci permettant d'obtenir un gradient électrochimique dont l'énergie est stockée par la suite par des liaison phosphates. Il suffit pour celŕ d'une membrane comportant des molécules perméables aux ions et dont l'une des faces est exposé ŕ un rayonnement: un gradient ionique, source potentielle d'énergie pour le métabolisme, se met alors spontanément en place. Un métabolisme - ancętre de la photosynthčse a donc pu apparaître trčs rapidement.
Toutefois,
la disponibilité
des ions phosphates dans le milieu réactionnel
constitué
par la Terre primitive est problématique (Gulick, 1955) : En
milieu aqueux, les ions phosphates réagissent avec le
calcium
pour former de l'apatite, un minéral quasiment insoluble.
Des
1972, Schwartz avait proposé que la formation du coeur
terrestre
se soit accompagnée d'un dégagement massif de
phosphates,
phosphites et acide phosphoriques obtenus par réduction
ŕ
haute température. Pour résoudre ce
problčme,
Glindeman & al. (1999) ont obtenu expérimentalement
la
formation de phosphites ŕ partir d'apatite, grace
ŕ des
décharges électriques. Ils proposent que les
éclairs accompagnant les nuages
dégagés par
des éruptions volcaniques ait pu contribuer ŕ la
formation de phosphates incorporables dans les molécules
prébiotiques, et ce dans plusieurs types
d'atmosphčres
possibles (N2 avec de 0 ŕ 10 % de méthane).
L'andésite, dégagée sous forme
pulvérulente
lors d'une éruption, contient ainsi en moyenne 0,3
% de P2O5 qui
peiut ętre soumis ŕ l'action des
éclairs. Le taux de production de ces
phosphites est estimé par Glindemann
ŕ 1.3×10−9 mol cm−2
yr−1, ce qui
est un taux présentant un large excčs par rapport
ŕ celui de formation
volcanique d'un autre précurseur prébiotique, le
HCN.
Les
phosphites, mille fois plus
solubles que le phosphate de Ca, conduisent ŕ la formation
d'acides
phosphoniques lorsqu'ils sont en présence de
molécules organiques et
d'unb rayonnement UV. Ces acides ont déjŕ
été mis en évidence dans des
météorites, ce qui démontre leur
formation en milieu spatiel et une
possible contribution extraterrestre ŕ l'enrichissement
enacides
phosphoniques de l'environnement primitif terrestre (Cooper et al.,
1992), et auraient pu constituer une source de composés
phosphatés pour
l'élaboration des nucléotides ou de leurs
ancętres.
On ne peut exclure également ŕ mon sens une synthčse des polymčres en milieu sec, avec apport continu d'énergie résultant du rayonnement stellaire.
Un autre mécanisme peut ętre invoqué comme source d'énergie: il s'agit de la formation de pyrite ŕ partir du sulfure d'hydrogčne, sous haute température et pression (Wachtershauser, 1988).
Dans les poussičres interplanétaires
Dans
les régions de
température assez élevée, des
molécules
prébiotiques se forment ŕ la surface de
minéraux
tels que les argiles et les sulfures. Ces réactions se
font dans une gangue de glace, dans un milieu semi solide. Elles
aboutissent ŕ la formation d'acides aminés et,
sans
doute, d'acides nucléiques. Dans les nuages
moléculaires,
il est possible d'obtenir des liaisons peptidiques (Wincel &
al., 2000) mettant en jeu
des intermédiaires
protonnés. Ces réactions en phase gazeuse ouvrent
la voie ŕ la constitution de peptides spatiaux qui peuvent
atteindre en grande quantité et sans dommage la surface
terrestre (Maurette, 1998).
Dans
l'atmosphčre des plančtes joviennes
|
Certains auteurs considčrent que la probabilité d'apparition de la vie est si faible que celle ci ne peut exister que sur Terre. Outre que la "diaspora bactérienne" met ŕ mal une telle conception, d'autres arguments peuvent ętre invoqués pour justifier une "exception Terrestre". Réglons leurs compte avant de poursuivre:
|
On se retrouve ŕ présent avec des "ARN" et des peptides adsorbés sur des surfaces minérales. Dés le départ, ARN et peptides sont liés. En effet, les peptides stabilisent les ARN formés. Comme la formation de ces molécules est orientée par la géométrie de catalyseurs minéraux, la croissance des cristaux de ces męmes minéraux, reproduisant leur répartition de charges ainsi que leur forme, fournit une multiplication des lieux de synthčse: la reproduction n'est alors que minérale, l'organique venant en surplus. En męme temps que ces synthčse minérales, le milieu liquide est le sičge d'un enrichissement en composés susceptibles de former des membranes. Nous allons bientôt les retrouver...