Une chimie avant la vie
Derničre mise ŕ jour: 21/08/2009
L'oxygčne,
un poison
indispensable.
Nous vivons dans
un monde dont
l'atmosphčre contient 20% de dioxygčne. Cependant, ce
gaz est corrosif et les ętres vivants s'en protčgent par
des dispositifs variés. Des enzymes comme la cathalase, la
peroxydase ou la superoxyde dysmutase permettent au
métabolisme de résister ŕ l'action
délétčre de l'oxygčne et de ses
dérivés.
Pour les
bactéries anaérobies
qui ne disposent pas de ces protections, l'oxygčne est
toujours un poison mortel. Sans oxygčne, pas d'ozone pour nous
protéger des UV solaires qui arrivaient directement au sol,
apportant l'énergie nécessaire aux synthčse
chimiques ŕ partir des éléments minéraux
disponibles: H2O, le CO2 et NH3.
De nombreux
indices montrent que
l'oxygčne était absent au début de la
vie:
- des dépôt d'uraninite UO2
datant de cette époque sont constitués. Ors, l'uraninite
est dégradée en U3O8 soluble si la teneur en O2 est
supérieure ŕ 1%
- Des couches rouge d'hématite Fe2O3
nous indiquent une teneur croissante en oxygčne avec le temps.
Il est ŕ noter que de tels dépôts d'hématite
ont été identifiés sur Mars, renforçant
encore l'hypothčse selon laquelle Mars et la Terre ont
évolué de concert aprčs leur formation.
- Des formations de fer rubanné se
sont déposées lorsque la teneur en O2 a augmenté,
transformant le Fe2+ soluble en Fe3O4 insoluble qui a
précipité avec de la silice
- Le métabolisme des ętres
vivant garde la trace de cette époque: toutes les
réactions chimiques de base se déroulent en l'absence
d'oxygčne. L'étude du métabolisme est importante
pour définir les origines de la vie: c'est un fossile
moléculaire gardant intact des indices précieux sur
l'aube de la vie...
ainsi, les
réactions
énergétiques qui permettent aux ętres vivant de
récupérer de l'énergie ŕ partir des
molécules de leur alimentation sont de 3 type
différents:
- la fermentation est le moyen le plus
ancien, de nombreuses cellules privées d'oxygčne
doivent y recourir, comme les cellules musculaires par exemple. C'est
la dégradation d'une molécule de glucose en 2
molécules de pyruvate. Celui ci est ensuite
dégradé en acide lactique ou en alcool éthylique,
mais constitue toujours un déchet. Chaque molécule de
glucose dégradée permet de mettre en réserve deux
"unités énergétiques" (UE) constitués par
des liaisons phosphates riches en énergie.
- la respiration utilise l'oxygčne
pour dégrader le pyruvate sous forme de CO2 et H2O. 36 UE sont
récupérées pour 1 molécule de glucose
dégradée! C'est ainsi que fonctionnent les cellules de
notre organisme. Si elles sont privées d'oxygčne, elles
"retombent en enfance" et effectuent des fermentations.
- la photosynthčse est
utilisée par les végétaux et permet de construire
des molécules telles que le glucose ŕ partir du CO2 et de
l'énergie lumineuse.
Dans
l'expérience de Miller, des
décharges électriques dans des mélanges de
H2,CH4,NH3 et
H2O
donnent des aldéhydes, des acides
carboxyliques et des acides aminés. Les męmes
résultats sont obtenus avec un mélange gazeux contenant
du CO, du CO2 et de l' N2 (si le milieu
réactionnel ne contient pas d'oxygčne).
Comment
sont apparus les
molécules du vivant ?
Sur la
centaine
d'atomes que l'on trouve
dans l'univers, 24 sont nécessaires aux molécules
constituants les ętres vivants. Les molécules
caractéristiques des ętres vivants sont de trčs
grandes tailles: on parle de macromolécules. Elles sont
formées de l'enchainement de molécules plus petites,
les monomčres. En simplifiant beaucoup, on peut distinguer 2
grands groupes de macromolécules caractérisant le
vivant:
- les protéines qui sont des
enchaînements d'acides aminés constituent les structures
des cellules. Dans les cellules, on trouve seulement 20 acides
aminés différents
- l'ADN et l'ARN contiennent et transmettent
les informations qui permettent la reproduction et le fonctionnement
des cellules. Ils sont formés de l'enchaînement d'acides
nucléiques. Il existe 5 acides nucléiques utilisés
par les ętres vivants. L'ARN est capable de se répliquer
lui męme et fut sans doute le premier acide nucléique des
ętres vivants. Pour le fabriquer, il faut disposer d'un sucre, le
ribose, de phosphate et de 4 bases différentes.
Longtemps les
chercheurs se sont
demandés qui des enzymes ou des acides nucléiques
étaient apparus en premier: ces deux familles de
molécules semblaient indispensable l'une ŕ l'autre,
mais en 1986 T. Cech montre que
l'ARN peut ętre son propre enzyme:
l'ARN semble ętre la premičre
macromolécule biologique apparue sur Terre. L'ARN
possčde de par sa structure une activité sur les
protéines. De nos jours, celle ci s'exerce dans des
assemblages complexes, les ribosomes, dans lesquels l'ARN catalyse la
formation de liaison peptidiques entre acides aminés. D'autres
ARN peuvent découper des protéines. De plus, les
molécules d'ARN possčdent une certaine capacité
d'évolution en réponse ŕ leur environnement:
l'ARN peut devenir résistant ŕ la ribonucléase
qui, normalement, le clive.
L'étape
qui sépare la
synthčse prébiotique des Acides Aminés (AA) et
des Acides Nucléiques (AN) des premičres
bactéries s'étale sur 500 millions d'années.
Aprčs la formation de la plančte, plusieurs
étapes ont du se succéder:
1-
synthčse des
monoméres biologiques (sucres, AA et AN)
Pour obtenir
les
molécules
indispensables au ętres vivants, il faut pouvoir trouver une
voie de synthčse minérale des AA, des sucres, des
phosphates, des bases organiques (pour les AN), des lipides (pour les
membranes) et quelques autres molécules comme les
flavines.
Dans
l'expérience de Miller, on
obtient des acides aminés tels que la Leucine, l'isoleucine,
la Thréonine, l'Asparagine, la lysine, la phenylalanine et la
tyrosine mais aussi des acides aminés et des isomčres
inconnus dans les ętre vivants: la sarcosine et la b alanine
par exemple. De plus, les 20 AA utilisés par les ętres
vivant ont tous la męme symétrie spatiale, dite
L (sauf
certaines paroi bactériennes et
quelques molécules de défense). Or, les
synthčses prébiotiques conduisent autant aux formes L
qu'aux formes D. Pourquoi donc seuls les AA de symétrie L sont
ils utilisés par les ętres vivants ? Il est possible que
le hasard soit ŕ l'origine de cette sélection, bien que
de nombreuses propositions ont été avancées,
faisant appel, entre autre aux forces de Coriolis résultant de
la rotation terrestre (qui ŕ l'époque était un
peu plus rapide que de nos jours, la lune ayant depuis freinée
la rotation terrestre).
On peut
également se demander
pourquoi seulement 20 AA entrent dans le code génétique
alors que davantage d'AA différents étaient disponibles
? Il est probable qu'ŕ l'origine, le code
génétique comportait moins de 20 AA, beaucoup d'entre eux
dérivant des autres. Au fil de l'évolutiion, de nouveaux
acides aminés ont été intégrés dans
les protéines, mais les organismes primitifs ne comportaient
peut ętre qu'une quinzaine d'AA différents, voire moins.
Il y eu
probablement, au début, des sélections aléatoires
basées sur les richesses en AA des différents milieux,
et pas mal d'essais disparus sans laisser de traces...
Les
monomčres ont commencés
ŕ se former en milieu gazeux, dans l'atmosphčre. Celle
ci était d'ailleurs enrichie en molécules organiques
par des molécules d'origine spatiale: les gaz interstellaire
et la poussičre contiennent de l'eau, de l'NH3, du
formaldéhyde, du méthane...
Le formaldéhyde a aussi pu se former dans l'atmosphčre
primitive par photochimie (Pinto & al., 1980).
|
Une
centaine de molécules diverses sont aujourd'hui connues dans
l'espace interstellaire, dont certaines comportant plusieurs dizaines
ŕ plusieurs centaines d'atomes de carbone.
Les
comčtes, trčs nombreuses aprčs la formation du
systčme solaire, ont également été une
source de HCN, CO2, formaldéhyde, méthanol, CO, NH3 et H2S mais
également d'eau pour l'atmosphčre primitive.
Les
premičres molécules intervenant sont le HCN, le
cyanogčne C2N2 ainsi que des aldéhydes.
Les AA se
sont formés ŕ partir de ces précurseurs, par la
synthčse dite de Strecker. Des hydroxyacides peuvent se former
de la męme façon, comme l'acide glycolique, le glycocolle,
l'acide lactique oů l'alanine.
Les
sources d'énergie disponibles ne sont pas que les UV, au
demeurant pas toujours efficace pour tous les gaz (ils sont surtout
absorbés par H2S et NH3). Les éclairs, la radioactivité ainsi
que les ondes de choc des impacts météoritiques et
cométaires ont pu apporter l'énergie nécessaire
aux synthčses prébiotiques. On peut également
noter que la présence d'ions Zn favorise la
polymérisation des AA en peptides. Des minéraux apportant
cet ion ont aidé ŕ l'éllaboration des premiers
peptides...
Les
éclairs offraient l'avantage de fournir une grande
quantité d'énergie ŕ des endroits bien
localisés.
Ci
contre: une voie de synthčse des acides aminés se
produisant dans deux milieux: dans l'atmosphčre jusqu'au stade
aminonitrile, puis dans les océans ou en milieu liquide ensuite.
|
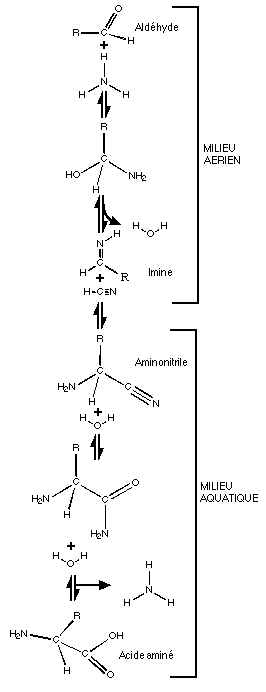
|
- Les sucres peuvent ętre obtenus par
condensations formolique, certaines bases peuvent dériver de
l'acide cyanhydrique (bases puriques: adénine et guanine)
grâce ŕ un intermédiaire, le diaminomaléodinitrile. La synthčse des autres bases (pyrimidiques:
l'uracile, la thymine et la cytosine) est bien plus
problématique.
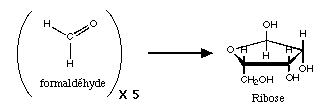
condensation
formolique aboutissant
au ribose
Il
faut toutefois remarquer que la synthčse de ribose "seul" est
problématique. Le rendement des réactions envisagées est mauvais, et
les conditions réactionelles spéciales. De plus, l'addition du ribose
sur les bases est encore un mystčre: le mécanisme est inconnu pour les
purines, et n'a été obtenu avec des pyrimidines qu'avec des molécules
de cette famille qui ne sont pas les bases des acides nucléiques
actuels.
synthčse
des Acides Nucléiques:
La
condensation de l'adénine et du ribose donne de
l'adénosine, un nucléotide qui en se liant ŕ 3
groupes phosphate donne de l'Adénosine Tri Phosphate, toujours
utilisé pour transférer l'énergie dans les
ętres vivants.
Pourquoi
utiliser l'ATP et non pas la Guanine Tri Phosphate (GTP), la Cytosine
Tri Phosphate (CTP) ou l' Uracyle Tri Phosphate (UTP)? La
synthčse de l'adénine se révélant plus
facile, peut ętre cette molécule fut elle tout simplement
celle qui était le plus aisément disponible.
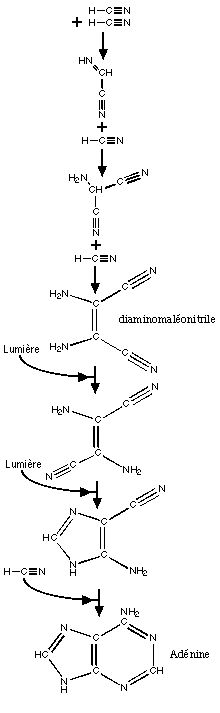
Ci
contre: voie de synthčse prébiotique de
l'adénine. Le rendement de cette réaction est cependant
trés
faible.
La "voie des purines"
L'examen du
métabolisme commun ŕ toutes les cellules (eucaryotes,
procaryotes...) donnent quelques indices montrant que les bases
puriques (A et G) ont probablement été les premiers
nucléotides utilisés par le vivant (hypothčse personelle).
En effet, la synthčse des purines présente nombre de
caractčres indiquant qu'il s'agit d'un mécanisme plus
fondamental, plus basique que celui des pyrimidines. Par exemple, la
fabrication de pyrimidines nécéssite l'existence
préalable de molécules basées sur
l'adénine, alors que le contraire n'est pas vrai.
Le tableau ci dessous met en lumičre ces
différences:
Molécules
|
Purines
|
pyrimidines
|
précurseurs de la synthčse
|
ŕ partir de
molécules simples: CO2, glycocolle, glutamine, Acide aspartique,
acide tetrahydrofolique.
La néosynthčse est plus importante que la
récupération d'origine alimentaire.
|
glutamine
nécessite l'intervention du NAD (donc la présence
d'adénine!)
|
mode de synthčse
|
Le ribose constitue la base sur
laquelle le cycle est progressivement édifié.
|
le cycle est
synthétisé puis fixé au ribose.
|
cofacteurs de la synthčse
|
minéraux:
2 Mg 2+, K+
|
Organique:
NAD
|
lieu de synthčse
|
Cytoplasme puis interconvertions
en
mitochondries |
Cytoplasme
|
"matériel"
moléculaire pour la synthčse
|
IMP, 9 ATP (donc de
l'adénine!).
10 enzymes
2 coenzymes
(toutes cellules sauf lymphocytes)
|
la conversion U--->C
nécéssite de l'ATP, donc... de l'adénine!
|
Ces
données plaident en faveur d'une utilisation
préférentielle des purines au début du code
génétique: le premier code pourrait avoir reposé
sur un acide nucléique formé d'un simple brin de purines...
Synthčse des pyrimidines (cytosine et uracile)
ILa
cytosine peut ętre obtenue dans les expériences type Miller, ŕ partir
de cyano-acetylene et de cyanate obtenus par décharge électrique dans
une atmoisphčre contenant un mélange de méthane et d'azote. Toutefois,
Miller lui męme fait remarquer que " this
reaction requires relatively high concentrations of cyanate (>0.1
M), which are unlikely to occur in aqueous media as cyanate is
hydrolysed rapidly to CO2 and NH3. " (cette réaction necessite
des concentrations relativement élevées en cyanate (> 0,1 mol/L),
qui ne peuvent ętre obtenues en milieu aqueux oů le cyanate est
rapidement hydrolysé en CO2 et NH3) - Michael P. Robertson, Stanley L. Miller. An efficient prebiotic synthesis of cytosine and uracil. Nature 375, 772-774 (29 Juin 1995) . Ces męmes auteurs proposent alors une voie de synthčse basée sur le cyanoacetaldehyde provenant de l'hydrolyse du cyanoacetylene, réagissant avec de l'urée.

Si cette derničre molécule est présente en abondance (en bord de lacs en cours d'évaporation, par exemple), alors le cyanoacetaldehyde
conduit ŕ la formation de cytosine avec un rendement compris entre 30
et 50%. L'uracile peut se former par hydrolyse de cette molécule.
Autre
problčme ŕ résoudre: on trouve aujourd'hui dans l'ARN une liaison
bęta 1'
alors que des alpha et bęta 1', 2' et 3' étaient aussi
possibles et tout aussi probables. Pourquoi cette liaison ŕ
t'elle été selectionnée par l'évolution ?
Il semble que des ions minéraux puissent expliquer cette
sélectivité: de nos jours encore, la synthčse
d'ADN et d'ARN est sous la dépendance des ions Mg et surtout Zn
qui orientent le sens des synthčses. Voici encore un indice de
l'implication des minéraux dans l'origine de la vie.
Nous
savons que des molécules de synthčse peuvent catalyser
leur reproduction, évoluer et se disputer les ressources de leur
milieu. C'est le cas de l'ARNI (adénine ribose naphtalčne
imide) qui peut se répliquer ou s'associer avec elle męme,
ou de la diaminotriazine xanthčne thymine (DIXT). Ces
molécules comprenant des
acides nucléiques peuvent
d'ailleurs se lier et engendrer par recombinaison d'autres
molécules douées d'autoreplication ou non. (J.Rebek
- Pour La
Science 203 - sept 1994).
Pour
qu'une molécule soit capable de s'autorépliquer, sa forme est
primordiale ainsi que la répartition des forces exercées
par les différents groupements moléculaires qui la
compose. Ces forces peuvent orienter
et attirer les molécules de base,
et la forme de la molécule permettre le rapprochement des diverses molécules afin de
faciliter leur réaction: c'est
le mode de fonctionnement des enzymes de nos cellules.
2
-
La
polymérisation des monomčres
Pour
enchaîner les différents
monomčres, il faut réaliser des réactions de
polymérisation. Pour cela, il faut lier deux molécules
en leur enlevant une molécule d'eau alors que dans un milieu
aqueux, c'est la dépolymérisation (hydrolyse) qui est
favorisée (empęchant
l'allongement des polymčres
protéiques ou nucléiques). Pour que cela soit possible, il faut
disposer de nucléotides "activés", c'est ŕ dire comprenant un
groupement phosphate. Mais comment relier ll'improbable ribose aux
bases ?
Une des façons de court-circuiter ce problčme est d'envisager une synthčse directe ŕ partie d'un précurseur de la base et du ribose (Matthew W. Powner, Béatrice Gerland, John D. Sutherland. Synthesis of activated pyrimidine ribonucleotides in prebiotically plausible conditions. Nature 459, 239-242 - 14 Mai 2009).
Pour cela, un ensemble de molécules prébiotiques simples, ŕ la présence
probable (cyanamide, cyanoacetylene, glycolaldehyde et glyceraldehyde)
ainsi que des phosphates minéraux réagissent, en présence d'azote et au
cours de cycles de réchauffement/refroidissement (simulant une
évaporation suivie de pluies, dans pendant une semaine pour
l'expérience réalisée par l'équipe pré-citée) pour former
du 2-amino-oxazole (des
mécanismes réactionnels conduisant ŕ cette molécule sont
détaillés sur l'excellent site, en anglais, du Dr Henry Jakubowski, de
l'Université St John).
Ce précurseur réagit avec le
glycéraldéhyde pour conduire ŕ un ensemble ribose et cytosine. Le
phosphate, en présence d'UV aboutit ensuite ŕ une synthčse de
ribocytidine phosphate, certains nucléotides formant aussi
de l'acide uridylique. Outre son rôle de réactif, le phosphate, indispensable dčs le début de la réaction,
régule ces derničres en jouant le rôle de tampon ainsi que de catalyseur. On obtient
"finalement", en quatres étapes, deux ribonucléotides phosphates: la
cytidine et l'uracyle.
Les auteurs font remarquer qu'il est
important de partir d'un mélange réactionnel comportant des réactifs
bombreux, et de ne pas ęytre tenté de "rajouter au bon moment" le
réactif qui "débloque la situation", ce genre de procédé étant trčs peu
probable dans les réelles conditions prébiotiques...
Il a donc fallu
que le milieu favorise la
polymérisation. Pour cela, il était nécessaire
de concentrer les réactifs ou d' enlever l'eau, et de coupler
la réaction ŕ une autre qui libérerait de
l'énergie. En effet,
l'allongement d'un polymčre
nécessite de l'énergie, qui doit ętre fournie par
couplage avec d'autres réactions chimiques par exemple, alors
que l'hydrolyse libčre de l'énergie, ce qui lui permet
de se produire spontanément. Il a donc fallu que la
polymérisation soit couplée
ŕ une autre réaction lui fournissant l'énergie
dont elle a besoin.
L'indispensable
mécanisme de
couplage énergétique
Certains
composés comme le
carbodiimide (-N=C=N-) fournissent assez d'énergie pour une
polymérisation. D'autres agents de couplage sont le
cyanogčne, le cyanamide, les cyanoacétylčne et
le diaminomaléodinitrile (leur énergie est
stockée dans une triple liaison CC ou CN). Pourquoi les agents
de couplage ne réagissaient ils pas plutôt avec l'eau
qu'avec les monomčres présents? La liaison entre les
monomčres et des ions phosphates ont pu favoriser ce couplage
au détriment de l'eau. Par cette voie, on a obtenu des
synthčses de polyphosphates (AN ). Les polyphosphates sont
utilisés encore de nos jours par certaines bactéries
qui stockent leur énergie dans des corpuscules ŕ
polyphosphates dans leur cytoplasme.
Enlever l'eau
favorise également les
mécanismes de polymérisation:
- L'adsorption sur des minéraux
argileux du type montmorillonite permet d'offrir une grande surface
réactionelle aux molécules qui sont d'ailleurs
orientées pars les forces électrostatiques s'exercant au
niveau moléculaires entre les différents feuillets de ces
minéraux, le tout en limitant la quantité d'eau
présente.
- Le mélange réactionnel peut
ętre concentré par évaporation ou
congélation.
- Des réactions de chimie
sčche sont possibles: S.Fox a montré que des AA sec
polymérisent ŕ 130°C . On obtient alors des
"thermo-protéinoďdes", des molécules qui seraient
les ancętres des protéines. Avec des polyphosphates, la
polymérisation ŕ sec se produit ŕ 60°C (On
peut ainsi aboutir ŕ des polymčres de 200 AA environ)
3 -
Ségrégation
dans des milieux protégés (membranes) des
polymčres obtenus
Les
polymčres tels que AN et
protéines en solution ont tendance ŕ former des
gouttelettes nommées coacervats. Ces gouttelettes sont plus ou
moins stables et peuvent concentrer des molécules en fonction
de leur solubilité.
On peut
également envisager la
formation de membranes protéiques (comme pour les capsides de
virus).
4 -
Développement d'un
mécanisme de reproduction
Ce
mécanisme devrait se baser sur
une polymérisation autocatalytique. Il est probable que les
premičres molécules autoréplicantes
étaient trčs simples, comportant peu de
monomčres. Leur réplication en fonction des apports du
milieu faisant augmenter le volume des membranes les contenant,
celles ci finissent par éclater en en formant de plus
petites.
L'environnement
dans lequel la vie est
apparue était trčs différent du notre, surtout
en ce qui concerne la température et la composition de
l'atmosphčre. Les débats sont encore vif en ce qui
concerne la température ŕ l'époque
prébiotique: alors qu'une origine chaude semblait se
dégager, l'étude des molécules d'ARN des
organismes actuels montrent que les séquences
sélectionnées ne sont pas celles qui résistent
le plus ŕ la chaleur. Par contre, il ne fait aucun doute que
l'atmosphčre primitive était dépourvue
d'oxygčne: ce gaz est une signature de la vie
végétale.
Dés son
apparition, la vie
détruisit les conditions de son éclosion. a partir du
CO2 atmosphérique, les premiers organismes
photosynthétiques ont formé du glucose en dissociant
H2O
avec l'énergie lumineuse, en
libérant O2 comme produit de la
réaction. Le milieu n'est pas un ensemble éternel dont
la composition est fixé et doit ŕ toute force
ętre préservée: pour les premičres
cellules, l'oxygčne était un polluant mortel! Il est
logique que le milieu évolue, et que nous contrôlions
cette évolution...
Références
Lire aussi en français:
- L'origine de
la vie sur la
Terre - L. Orgel - Pour La Science 206, décembre 1994
80-88
- L'origine de la vie - R. Shapiro (intéressant par son approche critique et constructive) - coll. Champs; Flammarion
- Les origines cosmiques de la vie - Delsemme (des intuitions pleinement confirmées) - coll. Champs, Flammarion