Le monde des polymères
Deux conceptions
différentes
se font jour quant aux premiers êtres vivants: 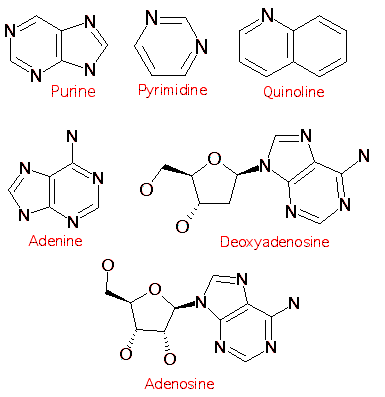 certains suggèrent un
système autoréplicatif, comportant quelques acides
nucléiques, alors que d'autres font remonter le premier
être vivant à l'apparition du premier échelon
du métabolisme: la fixation du carbone. Ces deux conceptions
ne s'excluent pas mutuellement de prime abord, elle ne différent
qu'au plan temporel et, surtout, au niveau des conditions initiales
qui sont supposées. L'étude de ces dernières
semble bien favoriser, sur la Terre primitive, l'option tirant
partie de fortes températures et pressions. La découverte
récente des nanobes laisse également entendre une
ancienne et forte liaison entre le monde minéral et organique.
Dans cette optique, et bien que je considère que l'essentiel
des synthèses ai pu se produire ailleurs que sur la Terre
primitive, le développement des premiers métabolismes
est une condition nécessaire et déterminante à
l'apparition de la vie. Nous l'intégrerons donc à
notre approche globale.
certains suggèrent un
système autoréplicatif, comportant quelques acides
nucléiques, alors que d'autres font remonter le premier
être vivant à l'apparition du premier échelon
du métabolisme: la fixation du carbone. Ces deux conceptions
ne s'excluent pas mutuellement de prime abord, elle ne différent
qu'au plan temporel et, surtout, au niveau des conditions initiales
qui sont supposées. L'étude de ces dernières
semble bien favoriser, sur la Terre primitive, l'option tirant
partie de fortes températures et pressions. La découverte
récente des nanobes laisse également entendre une
ancienne et forte liaison entre le monde minéral et organique.
Dans cette optique, et bien que je considère que l'essentiel
des synthèses ai pu se produire ailleurs que sur la Terre
primitive, le développement des premiers métabolismes
est une condition nécessaire et déterminante à
l'apparition de la vie. Nous l'intégrerons donc à
notre approche globale.
Ci contre: 6 types de molécules utilisées par les êtres vivant. Leur élaboration par ajout d'éléments successifs sur une structure de base commune est ici patente.
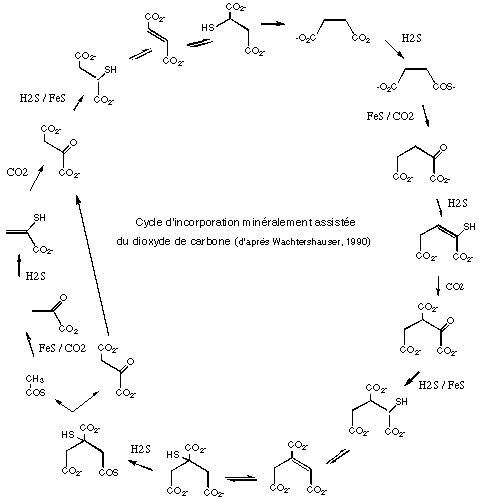
Notre hypothèse de départ sera que les premiers organismes pouvait être autotrophes. Les premiers métabolismes sont des réactions s'effectuent en deux dimension, sur des surfaces minérales. L'incorporation du carbone est assurée grâce au pouvoir réducteur (électrons) provenant de la formation de pyrite FeS2 (Wachtershauser, 1989). Nous avons vu que ce minéral est aussi disponible dans la poussière interstellaire. On obtient ainsi un cycle correspondant à une version archaïque du cycle de Krebs, permettant incorporation du carbone dans des molécules organiques et production d'énergie. Les intermédiaires réactionnels, électriquement chargés, peuvent rester lié de façon électrostatique à leur substrat minéral (Wachtershauser, 1994).
|
Expérimentalement,
il est possible
de former des liaisons peptidiques avec du CO et des sulfures
métalliques (Ni, Fe). 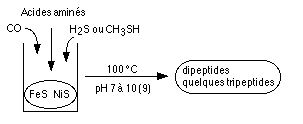 On obtient ainsi des
dipeptides,
voire quelques tripeptides (Huber & al., 1998). La
formation
de ces liaisons nécessite de l'énergie. Celle ci
peut être fournie par le cycle du CO, des diketopiperazines
et carbo-di-imides (Brack, 1993) ou du Cu en solution
salée
(Schwendinger & al., 1995). Cependant, le
mélange Cu/NaCl
semble avoir été rare dans les conditions primordiales
généralement invoquées, et les carbo-di-imides,eux
mêmes organiques, ainsi que les di-ketopiperazines semblent
se former principalement, la glycine n'étant qu'un à
coté de leur synthèse.
On obtient ainsi des
dipeptides,
voire quelques tripeptides (Huber & al., 1998). La
formation
de ces liaisons nécessite de l'énergie. Celle ci
peut être fournie par le cycle du CO, des diketopiperazines
et carbo-di-imides (Brack, 1993) ou du Cu en solution
salée
(Schwendinger & al., 1995). Cependant, le
mélange Cu/NaCl
semble avoir été rare dans les conditions primordiales
généralement invoquées, et les carbo-di-imides,eux
mêmes organiques, ainsi que les di-ketopiperazines semblent
se former principalement, la glycine n'étant qu'un à
coté de leur synthèse.
La fixation du CO2 peu également permettre la synthèse d'acide urique, lequel peut être considéré comme une purine très oxydée. Les premier acides nucléiques pouvant se former aurait alors été les ancêtres de purines. Cependant, les premières bases puriques, simplement liées à un support de trioses phosphates (Wachtershauser, 1994), n'aurait pas, au début tout du moins, contenu d'information: ce qui se reproduisait, c'était tout d'abord des cycles métaboliques dont la divergence était assurée par les phénomène de croissance du support minéral.
Les premières formes de vies auraient donc été des structures planes, fixant le carbone, croissant grâce à leur support minéral et non délimitées par une membrane. L'individualisation (séquestration) dans une membrane protéo-lipidique sera un stade suivant de l'évolution de ce probionte, suivie ensuite par la disparition du support minéral de type pyrite.
Il me semble
que le plus grand danger
pour ces organismes c'était, tout simplement, la dilution,
voire l'hydrolyse. Pourquoi ne s'en seraient ils pas
protégé
par une enveloppe fortement hydrophobe, capable de fixer des
éléments
minéraux afin d'accroître sa résistance
mécanique
? On se retrouverait alors en présence d'une pression de
sélection favorisant des organismes de type nanobes...
Dans les parois bactérienne Gram + actuelles existent
des molécules qui présentent des analogies surprenantes
avec les acides nucléiques: les acides teichoïques
(poly-glycerol-phosphates et poly-ribitol-phosphates nantis de
radicaux variés, liés à des peptidoglycanes).
Elles sont constituées, comme les A.N., d'une alternance de
sucres et de phosphate mais l'emplacement des bases est occupé
soit par un acide aminé soit par un sucre supplémentaire (Shapiro,
1998). Ces molécules sont des sites de fixation pour les
virus. Ces "échelles moléculaires" ont pu participer
à l'ellaboration des acides nucléiques modernes, matériaux d'abord structurels avant
de devenir fonctionnels (Bien
que le Pr. Shapiro dans son livre "l'origine de la vie", p 370 puis
378, mentionne cette notion, il ne la développe pas et ne voit
pas que le ribosome actuel, qu'il mentionne, correspond trait pour
trait à l'ancêtre des formes de vies qu'il invoque...).
Les parois bactériennes sont riches en molécules
exotiques pour les autres êtres vivants: on y trouve en effet des
acides aminés D et des peptidoglycanes, inconnus des eucaryotes.
L'intérêt
des purines
Nous avons vu que les premieres
molécules d'acides nucléiques formées ont sans
doute été des purines, ce que
nombre d'indices corroborent.
L'examen des voies de synthèse
actuelles des A.N. puriques fournit les indices suivants:
De nos jours, certains micro-organismes vivant dans des milieux pauvre en oxygène présentent un métabolisme lié aux minéraux (Stevens & al., 2000): A température élevée (60°C) et en milieu légèrement basique, l'eau réagit avec les silicates ferreux des basaltes, ce qui provoque la mise en solution d'ions Fe2+ et d'hydrogène. Les bactéries utilisent ce dernier pour leur métabolisme anaérobie. L'hydrogène sulfuré H2S est également utilisé de nos jours par des bactéries endosymbiotiques des vers tubicoles Riftia pachyptila, vivant à la sortie des "fumeurs noirs", des sources chaudes abyssales riches en composés chimiques élémentaires.
L'eau, un dangereux solvant pour les molécules prébiotiques...
La
polymérisation des acides
nucléiques actuel se traduit par la formation d'une liaison
phospho-diester. 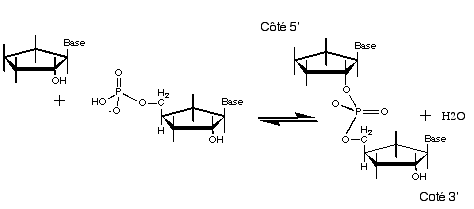 Le groupe OH du carbone 3' de la
partie "sucre"
d'un nucléotide forme une liaison avec le phosphate d'un
autre nucléotide. Cette réaction nécessite
de l'énergie, apportée actuellement par les liaisons
phosphates des nucléotides près à
polymériser.
Cette réaction produit, en outre, de l'eau. Comme elle
est réversible, elle a donc tendance, spontanément,
à aboutir à la destruction (hydrolyse) de
polymères
dans l'eau, avec production d'énergie... Il faut noter
qu'une chaîne simple d'acides nucléique est un poly-ester,
et que celui ci possède une orientation: une
extrémité
possède un groupe OH libre lié au carbone 3' alors
que l'autre bout de la chaîne n'est accessible que par un
groupe OH ou phosphate porté par la carbone 5'. Cette
orientation
est d'une grande importance dans les phénomènes
de réplication des acides nucléiques...
Le groupe OH du carbone 3' de la
partie "sucre"
d'un nucléotide forme une liaison avec le phosphate d'un
autre nucléotide. Cette réaction nécessite
de l'énergie, apportée actuellement par les liaisons
phosphates des nucléotides près à
polymériser.
Cette réaction produit, en outre, de l'eau. Comme elle
est réversible, elle a donc tendance, spontanément,
à aboutir à la destruction (hydrolyse) de
polymères
dans l'eau, avec production d'énergie... Il faut noter
qu'une chaîne simple d'acides nucléique est un poly-ester,
et que celui ci possède une orientation: une
extrémité
possède un groupe OH libre lié au carbone 3' alors
que l'autre bout de la chaîne n'est accessible que par un
groupe OH ou phosphate porté par la carbone 5'. Cette
orientation
est d'une grande importance dans les phénomènes
de réplication des acides nucléiques...
La polymérisation des acides
aminés présente des caractéristiques similaires:
production d'eau et énergie nécessaire. Pour former
cette liaison, la fonction amine d'un acide aminé réagit
avec la fonction "acide" d'un autre pour donner une
liaison CONH, avec élimination d'une molécule d'eau
Spontanément, en milieu aqueux, les protéines ont
donc tendance à se découper, et non à s'assembler.
On doit donc supposer que la formation des premières
molécules
s'est faite dans un milieu comportant assez peu d'eau.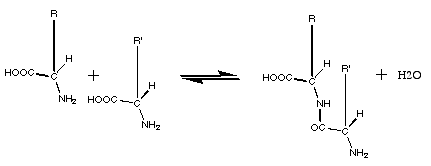
Les espaces
entre les feuillets
minéraux
des argiles et de certains cristaux sont particulièrement
indiqués pour cela. On doit également considérer
le rôle joué par des immersions et des emersions
successives du milieu réactionnel, causé par exemple
par les marées. La vie ne se serait alors pas
développée
dans la "soupe primitive" mais sur les cotés
de la soupière! Pourquoi ne pas pousser plus loin encore,
et considérer que, tout comme cela fut le cas pour
l'oxygène,
si l'eau est aujourd'hui indispensable à la vie, cela n'a
pas toujours été le cas, celle ci, au tout début
du moins, ayant pour principal problème de se protéger
des effets délétères de ce solvant universel
? On pourrait envisager un début de la vie "a sec",
donc possible même dans l'espace interstellaire, le "sec"
étant fournit en fait par le froid (jusqu'à
un certain point)!
Le fait que les réactions de polymérisation soient réversible, et dans le "mauvais" sens, en milieu aqueux, impose qu'il a fallu orienter ces synthèses par un apport d'énergie et une protection vis à vis de l'eau.
Parmi les polymères qui se sont formés avec les premiers acides nucléiques et peptides, on doit signaler des dérivés d'hydrocarbures, les terpènes. Ces molécules résultent de la polymérisation de l'isoprène (C5H8)n fois. L'isoprène CH2=C(CH3)-CH=CH2 se forme aisément, même dans le milieu interstellaire. Leur importance est grande car ils constituent la base des structures moléculaires constituant les premières membranes capables d'isoler les organismes de leur milieu.
Les fonctions des grosses molécules biologiques découlent de leur forme, et non, en fait, de leur composition chimique. Je m'en explique: la séquence d'une protéine conditionne son repliement, certes, mais toute séquence donnant une forme repliée identique donnera, au final, une fonction semblable ou similaire! Le repliement des protéines, pour prendre cet exemple, dépend de deux facteurs:
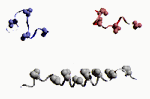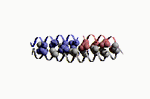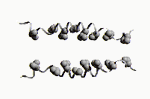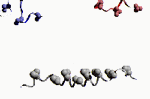 La
même situation se retrouve pour les acides nucléiques:
plus que leur séquence, c'est leur repliement, leur appariement,
leur structure spatiale qui est à l'origine de leur
activité:
les ARN de transfert agissent en fonction de leur forme, l'ADN
s'enroule, se surenroule et se déroule, exposant ou
protégeant
des séquences qui peuvent être transcrites ou non,
les ARNm se replient et se découpent... ces
phénomènes
sont intimement liés à la forme des molécules:
d'immenses séquences de l'ADN sont très variables,
non transcrites, sans utilité connue et les mutations peuvent
s'y accumuler par ce qu'elles ne perturbent pas le repliement
dans l'espace des macromolécules: la fonction découle
de la forme, laquelle dépend à la fois d'une partie
d'importance variable de la composition chimique et de l'environnement
de la molécule.
La
même situation se retrouve pour les acides nucléiques:
plus que leur séquence, c'est leur repliement, leur appariement,
leur structure spatiale qui est à l'origine de leur
activité:
les ARN de transfert agissent en fonction de leur forme, l'ADN
s'enroule, se surenroule et se déroule, exposant ou
protégeant
des séquences qui peuvent être transcrites ou non,
les ARNm se replient et se découpent... ces
phénomènes
sont intimement liés à la forme des molécules:
d'immenses séquences de l'ADN sont très variables,
non transcrites, sans utilité connue et les mutations peuvent
s'y accumuler par ce qu'elles ne perturbent pas le repliement
dans l'espace des macromolécules: la fonction découle
de la forme, laquelle dépend à la fois d'une partie
d'importance variable de la composition chimique et de l'environnement
de la molécule.
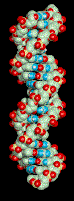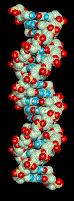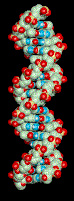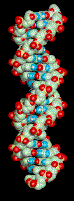
ci dessus: un modèle de molécule d'ADN sous lequel un modèle du peptide autoreplicatif obtenue par l'équipe de Saghatelian est représente (schéma NASA - Ghadiri MR )
Sans arrêt, nos molécules se tortillent, changent de conformation: l'ADN, par exemple, est bien moins une statique (un bloc de données) qu'une dynamique (une forme). L'ADN ne doit ses propriétés qu'à sa forme, qui n'est conditionnée qu'en partie par la séquence des éléments chimiques dont il est constitué (il peut d'ailleurs adopter plusieurs conformations selon l'environnement). Il en résulte qu'a l'échelon moléculaire, créer une forme revient à créer une fonction. Cette voie est expérimentalement explorée par les spécialiste de chimie supramoléculaire, qui tentent de créer des "enzymes de synthèse" et des molécules autoreplicatives simples (Rebek, 1994, Saghatelian, 2001).
 Nous
pouvons conclure de ce qui précède que la transmission
d'une information "génétique" revient
en fait à la transmission d'une forme particulière.
On peut en déduire qu'au début de la vie, ce qui
était transmis entre les générations d'êtres
vivant successif était davantage une forme qu'une structure
biochimique bien déterminée. Ce sont les minéraux
qui, une fois de plus, nous fournissent les premières formes
transmissibles susceptibles d'être considérées
comme une "information génétique" primitive.
Nous
pouvons conclure de ce qui précède que la transmission
d'une information "génétique" revient
en fait à la transmission d'une forme particulière.
On peut en déduire qu'au début de la vie, ce qui
était transmis entre les générations d'êtres
vivant successif était davantage une forme qu'une structure
biochimique bien déterminée. Ce sont les minéraux
qui, une fois de plus, nous fournissent les premières formes
transmissibles susceptibles d'être considérées
comme une "information génétique" primitive.
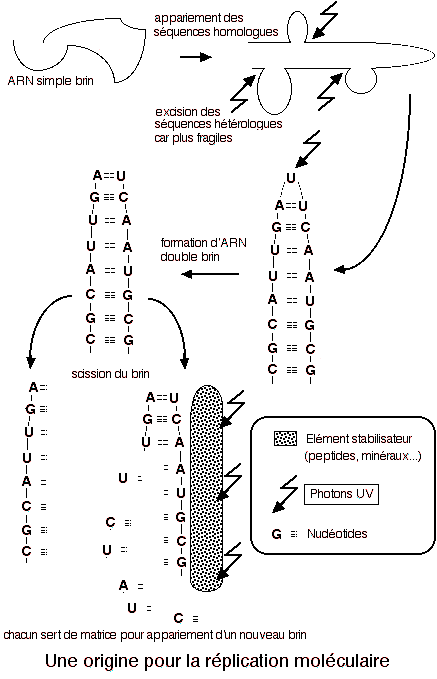
Le schéma ci contre montre
comment, dans le principe, a pu s'effectuer le passage de filaments
d'ARN simple brin à une molécule double brin capable
de faciliter sa réplication. On voit ici que dans un premier
temps les rayonnements UV ont été utiles à
l'évolution moléculaire. L'es éléments
stabilisateurs ont pu rester minéraux ou bien être
constitués de pigments primitifs à base de quinones,
capables de capter l'énergie des rayonnements pour la restituer
sous forme de liaisons chimiques rompues ou créées.
Un
point important mérite d'être précisé: outre
une reproduction des diverses molécules se recopiant avec des
fortunes diverses, il existait aussi des phénomènes de
coalescence de molécules différentes, de mélange
de "génomes" différents au sein d'une même roche.
En effet, l'information génétique primitive devait se
caractériser par une instabilité importante, les cassures
et recombinaison de molécules voisines devant être la
régle plutot que l'exception.