Notre scénario d'apparition de la vie faisant intervenir la poussière interstellaire, il implique une large distribution de cette dernière...
La vie a t'elle pu aussi
apparaître
sur une autre planète que la Terre ? Bien entendu, si des conditions similaires
s'y sont présentées. Mais contre ceux qui estiment
que la Terre est la seule planète habitée de l'univers,
on peut montrer que l'existence de la vie sur Terre implique
automatiquement la dispersion de cette même vie à
l'échelle du systéme solaire, voire d'une vaste
zone de notre galaxie.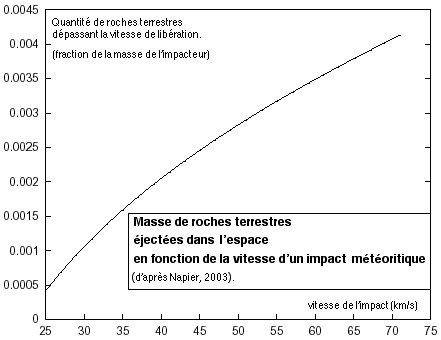
En effet, nos bactéries ont pu quitter notre propre planète depuis 4 milliards d'année à la faveur d'impacts météoritiques projetant dans le système solaire des roches contenant et abritant des micro-organismes. On compte un impact majeur du type Chicxulub tous les 100 MA environ sur notre planète. Chacun de ces impact est susceptible d'envoyer dans l'espace entre 10(9) (un milliard, Melosh, 1988) et 10(11) - 100 milliards (Napier, 2003, Wallis, 2003) de tonnes de matériel terrestre contaminé par des bactéries projetées bien vivantes dans l'espace. Cela signifie qu'en moyenne (bien que cette notion soit ici peu pertinente concernant des évènements majeurs si rares), notre planète expédie chaque année 20 tonnes de matériaux biologiquement contaminés dans le système solaire.
Quelle est la taille du domaine que j'apellerai "isobiologique", dans lequel ces matériaux ont pu ensemencer d'autres planètes, si un tel processus est possible ?
| Corps percutant la Terre |
|
|
| astéroides (>1 km) croisant notre orbite |
|
|
| comètes à courte période |
|
|
| comètes à longue période |
|
|
| comète type Halley |
|
|
Tours et détours dans le systéme solaire
Précisons le rôle de trois structures de notre systéme solaire qui influencent le destin des météores terrestres contaminés:
|
|
quantité de matériel terrestre contaminé reçu / million d'année (en tonnes) | temps du parcours de la terre vers l'objet (en millions d'années) |
| Terre (retour à l'envoyeur) |
|
|
| Mercure & Vénus |
|
|
| Mars |
|
|
| Jupiter |
4 millions |
100 |
| Ceinture de Kuyper |
|
|
|
Ors du système solaire: - expédition directe - via les corps de la ceinture de Kuyper |
|
3000 |
Nous pouvons calculer avec stupéfaction que nos bactéries voyageuses mettront 75000 ans pour parcourir 5 année lumière et atteindre l'étoile la plus proche. Pour nous cette durée est colossale, pour une bactérie, ce n'est rien... Des calculs effectués à l'université de l'Arizona (Melosh, 1985, Vickery, 1987) ont montrés que 25% des météores émis lors d'un choc sur la surface martienne ont été expédiés ors du système solaire par la force de gravité de Jupiter, prouvant ainsi que l'injection de roches dans le milieu interstellaire est une réalité.
|
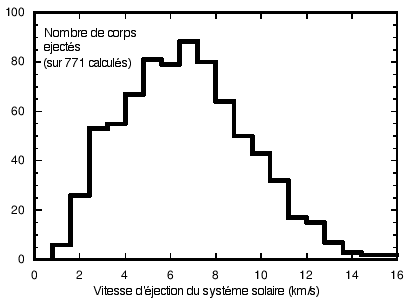
Ejection de roches d'origine terrestre du systéme solaire par Jupiter (d'après Melosh, 2003). Sur 5000 corps, 771 sont éjectés soit 15 %. La plupart des fragments ont une vitesse voisine de 5 km/s. (Avec l'aimable authorisation de Jay. Melosh, Regents Professor of Planetary Science, Lunar and Planetary Lab, University of Arizona, Tucson) |
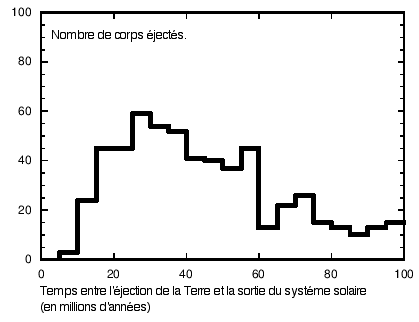
Temps de transit dans le systéme solaire avant éjection par Jupiter (d'après Melosh, 2003). Entre 20 et 40 millions d'années sont nécessaires avant éjection du systéme solaire. (Avec l'aimable authorisation de Jay. Melosh, Regents Professor of Planetary Science, Lunar and Planetary Lab, University of Arizona, Tucson) |
 |
 |
Cette injection est réalisée de plusieurs manières:
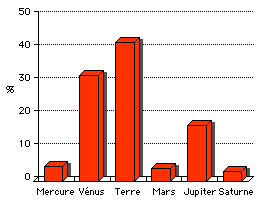 Ejections
directe de fragments: 20 %
des ejectas terrestres parviennent à l'orbite de Jupiter. Leur
vitesse est alors de quelques km/s, insuffisante pour sortir du
systéme solaire. Cependant, l'effet de fronde gravitationelle de
Jupiter expédie 1 à 2 % des ejectas Terrestres au
delà de Saturne (Melosh, 2003 - cf
schéma ci contre, réalisé d'après la
publication, montrant la destinée des roches terrestres
ejectées dans le systéme solaire). L'influence de Saturne et les effets
conjugés d'Uranus et de Neptune peuvent alors fortement
accélérer les roches et leur permettre de s'évader
du systéme solaire. Chaque année, une quinzaine de corps
de taille métrique doivent ainsi sortir du systéme
solaire dans lequel ils ont voyagés pendant une durée
moyenne de 50 millions d'années.
Ejections
directe de fragments: 20 %
des ejectas terrestres parviennent à l'orbite de Jupiter. Leur
vitesse est alors de quelques km/s, insuffisante pour sortir du
systéme solaire. Cependant, l'effet de fronde gravitationelle de
Jupiter expédie 1 à 2 % des ejectas Terrestres au
delà de Saturne (Melosh, 2003 - cf
schéma ci contre, réalisé d'après la
publication, montrant la destinée des roches terrestres
ejectées dans le systéme solaire). L'influence de Saturne et les effets
conjugés d'Uranus et de Neptune peuvent alors fortement
accélérer les roches et leur permettre de s'évader
du systéme solaire. Chaque année, une quinzaine de corps
de taille métrique doivent ainsi sortir du systéme
solaire dans lequel ils ont voyagés pendant une durée
moyenne de 50 millions d'années.Le long voyage
En 1,5 millions d'année, des bactéries voyageuses auront parcouru 100 années lumières, rencontrant plusieurs étoiles et tombant, éventuellement, sur leurs planètes... Cependant, les probabilités sont inflexibles et cette contamination directe très peu efficace: 1 météore d'origine terrestre serait capturé par une planète non solaire par milliard d'années, et les chances d'atteindre une planète "viable" (bien que ce qualificatif soit très imprécis) sont de 1 sur mille en 4,5 milliards d'années... En effet, la section de capture (surface que notre météore doit traverser pour être capturée par un autre systéme solaire) est très peu étendue: elle est voisine de 10(-11) AL2 (0,05 UA2 - Melosh, 2003), soit une surface 5 fois inférieure à celle couverte par l'orbite de Mercure pour des systémes planètaires comparable au notre (cette surface augmente un peu si le météore contaminé posséde une faible vitesse et si le systéme planétaire contient une planète bien plus grosse que Jupiter...)
Toutefois, il existe d'autres voies pouvant assurer la dispersion bactérienne au niveau galactique, résumées dans le tableau suivant:
|
|
|
|
| Apport direct | Un météore pénètre l'atmosphère d'une planète extrasolaire ou les conditions physiques permettent la reproduction des bactéries qu'elle contient. | apparemment négligeable pour les planètes de type terrestre (joviennes ?) |
| "Vaporisation" d'un noyau cométaire |
Les matériaux terrestre s'incorporent lentement aux noyaux cométaires de la ceinture de kuypers. Par la suite, ces noyaux voyagent et pénètrent d'autres systémes solaires qu'ils contaminent (0,02 % vont vers le centre d'un SS, sous forme cométaire...) au moyen de la chevelure cométaire et par les poussières émises (dès une distance de 5UA) Les
matériaux contaminés
peuvent être transférés par accrétion lente
dans les atmosphères planètaires ou par l'explosion de
noyaux cométaires dans l'atmosphère
(phénomènes observés sur Terre et sur Jupiter).
|
750 g de matériaux terrestres seraient tranférés/ système se formant dans une nébuleuse proche. |
| Contamination d'un nuage moléculaire | Les noyaux cométaires et astéroïdes transportent ors du systéme solaire 4 millions de t de matériaux tarrestres/ million d'année. Lorsque ces corps pénètrent un nuage moléculaire, les rayonnements stelaires importants rendent la comète active et elle se dégaze en 0,15 AL., donc contamine les marges seulement de ces régions. Certains auteurs (Wallis, 2003) considèrent qu'à cette occasion 10(21) µ-organismes peuvent être transfèrés.dans la nébuleuse. |
Le systéme solaire traverse un nuage moléculaire de plus de 1000 masses solaires tous les 220 MA. une nébuleuse serait contaminée tous les 15 MA, et ce à une distance inférieure à 300 AL. 1 t de matériel terrestre / système stellaire formé |
| Inclusion dans un disque protoplanétaire | Dans un nuage protoplanétaire, les particules émises par un noyau coimétaire sont protégée des rayonnements et inclus dans les planétoïdes en formation. Par la suite, ils contamineront les planètes présentes. | quelques Kg à quelques AL de distance |
On doit également prendre en compte que la formation des étoiles s'effectue dans une zone dense, possédant une population stellaire riche: les étoiles naissent en groupes, puis se séparent. Comme le soulignent Adams & Spergel (2005), la probabilité d’échange de matéirel biologique est fortement augmentée dans un amas stellaire où les étoiles sont à la fois proches et animées de faibles vitesses différentielles. Les simulations de ces chercheurs montrent que la probabilité de capture d'un élément minéral venant d'un autre systéme stellaiure varie entre 1/1000 et 1/1 million, selon le taux d'éjection et la taille de l'amas. Vu le grand nombre de roches éjectées d'un systéme stellaire, ces chercheurs concluent que toutes les étoiles d'un amas sont susceptibles de capturer des roches venant des autres systèmes stellaires de leur amas. Bien qu'une faible fraction des roches ejectée soit porteuse de formes de vies, la capture d'un de ces "vaisseaux habité" intervient entre 10000 et 16000 fois dans un amas. Les deux auteurs précédent ont alors calculé que, dans un amas, la contamination réussie d'une planète de type terrestre doit se produire entre 2 et 1000 fois. Toutefois, ils n'ont pas pris en compte la possibilité de contamination d'une atmosphère de planète jovienne, dont la probabilité est supérieure.
En 150 millions d'années, un météore contaminé peut couvrir 10000 années lumière, soit une bonne part de la galaxie où les étoiles de type solaire ne sont séparées, en moyenne, que de 8 AL... Or, plusieurs chercheurs (Cano, 1995; Vreeland, 2000) ont observé des bactéries capable de survivre jusqu'a 250 millions d'années au bas mot... mais le pourraient-elles dans l'espace ?
Un problème: résister aux radiations
Outre la poussière interstellaire, il nous faut prendre en compte les rayonnements qui parcourent l'espace: on y trouve des rayons UV (près des étoiles, mais absorbés par quelques mm de poussière), des rayonnements alpha, voire gamma pour les rayons cosmiques à l'origine encore fortement discutée, pour ne pas dire inconnue...
L'irradiation s'exprime en Gray (Gy). 1 gray correspond à l'absorption d'une joule d'énergie par 1 Kg de matière. A titre de comparaison, la norme européenne EN 552 considère que 25Kgy constituent une dose permettant de stériliser efficacement le matériel médical (à condition que celui ci soit propre, donc que la population bactérienne y soit déjà réduite!).
Certains auteurs considèrent que l'intensité des radiations dans le milieu spatial est telle qu'elle empêche toute vie sur les roches arrachées à notre planète. C'est faire peu de cas de la résistance des formes de vie bactériennes aux rayonnements ionisants et de la protection constituée par la roche qui enferme les micro-organismes. Il nous faut aussi compter avec les différences importantes de l'intensité du rayonnement en fonction de la distance au soleil: le problème n'est pas le même au voisinage de Mercure ou au vosinage de Pluton!
Vu l'intensité des radiations dans le systéme solaIre extérieur, Wallis (2003) évalue la durée de vie de bactéries sans protections particulières entre 1 et 10 MA. Rappellons que quelques cm de roches diminuent le flux de radiations solaires de 99,9 %, prolongant d'autant la survie des bactéries. L'intensité du rayonnement entre Mars et Jupiter est évalué à 0,3 KGy/ans, soit 100 Kgy en 300 ans (la bacterie Deinococcus radiodurans resiste à 10 Kgy administrés en un temps trés court!, alors qu'une éruption solaire déverse environ 3 Kgy à la distance de la Terre). Plusieurs travaux (Bresler & al., 1980; Kalinin & al., 1981; Pavlov & al., 2006) montrent qu'une alternance de périodes d'irradiation et de protection puet permettre de sélectionner des populations bactériennes et d'augmenter considérablement leur résistance aux radiations. L'équipe de A.K. Pavlov considère même que la radiorésistance de Deinococcus radiodurans ne peut s'expliquer que par l'origine en partie (les ancêtres de la bactérie ayant effectués un aller-retour Terre-Mars) Martienne de ce micro-organisme, la surface terrestre n'ayant pas permis l'émergence d'une telle résistance aux radiations. (Pavlov, 2006).
Il existe des mesures expérimentales qui permettent de soutenir ces affirmations. Des expériences menées en orbite terrestre ont montrée (Horneck & al., 1994) que des spores de bacillus subtilis résistaient pendant 6 ans au froid et au vide spatial. Pour ce qui est des radiations, une protection anti UV (poussières par exemple) assurait un taux de survie de 80 % (la rayonnement était de 5 Gy). Même sans aucune protection, 1 spore sur 10000 a résisté aux conditions spatiales. Ultérieurement, d'autres travaux (Rettberg & al., 2002) ont montré que la protection contre les UV solaires nécéssitait une bonne épaisseur de matériaux inorganique mais pouvait aussi être obtenu par le destruction des premières "couches" de bactéries, ces dernières se comportant alors comme un bouclier anti-UV efficace protegant les micro-organismes qu'elles recouvrent. Comment le concluent ces chercheurs: " a hypothetical interplanetary transfer of life by the transport of microorganisms inside rocks through the solar system cannot be excluded, but requires the shielding of a substantial mass of anorganic substances." De plus, même des organismes plus complexes que les bactéries, comme les lichens Rhizocarpon geographicum et Xanthoria elegans sont capable de survivre aux conditions spatiales: 90 % ont résistés à 15 j passés en orbite terrestre, au contact du vide, du froid et des radiations (Exp Foton M2, Space daily 9/11/2005 )
La zone isobiologique
Même si un météore parti de notre système
solaire il y a 3,5 milliard d'années a pu parcourir...
233000 années lumières soit deux fois le diamètre
de notre galaxie, il a subit l'influence des rayonnements cosmiques
mais aussi celui de la poussière interstellaire: le vide
interstellaire n'est pas tout à fait vide, et même
la très faible densité de matière qu'il abrite
peut suffire, sur de très longues distances, à
éroder
les météores. Un taux d'abrasion de 50 µm/pc
semble réaliste, ce qui signifie qu'aucun corps
centimétrique
ne pourrait franchir plus de 100 pc, soit environ 330 AL. Comme
tous les corps ne sont pas centimétriques, nous pouvons
estimer 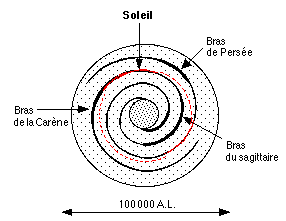 l'extension maximale
de la zone dans laquelle nos bactéries ont pu aissemmer
à un tore de 400 AL de rayon s'étendant sur
176000 AL! Ainsi, une
vaste région de notre galaxies peut avoir été
contaminées par des formes de vies microbiennes identiques:
l'échelon de base de la vie est le même dans cette
zone galactique de 88 milliards d'AL(3) renfermant plus de 2 milliards
d'étoiles
de type solaire !
l'extension maximale
de la zone dans laquelle nos bactéries ont pu aissemmer
à un tore de 400 AL de rayon s'étendant sur
176000 AL! Ainsi, une
vaste région de notre galaxies peut avoir été
contaminées par des formes de vies microbiennes identiques:
l'échelon de base de la vie est le même dans cette
zone galactique de 88 milliards d'AL(3) renfermant plus de 2 milliards
d'étoiles
de type solaire !
"Ma" zone isobiologique se trouve contenue dans la zone habitable galactique (GHZ) annulaire, comprise entre 23000 et 29000 AL du centre galactique définie par Lineweaver & al. (2004)
Sur ce schéma de notre galaxie, la zone isobiologique (rouge) montre sa forme annulaire. Parmi les zones les plus susceptibles d'avoir été contaminées, on trouve une partie du bras de la Carène ainsi qu'une région du bras de Persée. Ces bras faisant 200 AL d'épaisseur, on peut les considérer comme étant totalement contaminables.
On peut toutefois
remarquer que la contamination de la Terre primitive elle même
par un météore porteur de vie il y a 4 milliards
d'années est extrémement improbable (Davies, 2003)
car ce dernier proviendrait d'une époque où les
éléments lourds, moins nombreux, auraient eu moins de
chance de conduire à la vie. 
On doit donc en déduire qu'il existe probablement dans la galaxie une vaste zone "isobiologique" dans laquelle les micro-organismes sont, fondamentalement, similaires. Est il besoin de signaler que chaque monde sur lequel la vie se développe devient lui aussi une source potentielle de contamination de la galaxie? Une réaction en chaine s'initie donc inévitablement. D'où notre conviction, notre "théorème": pourqu'une galaxie contienne de la vie sur de nombreuses planètes différentes, il faut et il suffit que la vie y soit apparue sur une planète au moins"...
Le
point de départ
de l'histoire biologique qui, sur Terre, a aboutit à
la vie intelligente, est donc probablement le même sur de
nombreuses planètes de notre galaxie. A partir de ce point, bien
des divergences sont possibles. Est ce que cela conduit
nécessairement à l'apparition de l'intelligence
? Cela peut se discuter, car l'évolution est fonction de
l'environnement. Il est des planètes ou la vie reste
microbienne,
car les conditions physiques ne permettent pas l'apparition
d'organismes
plus massifs (je pense aux planètes joviennes). Il en est
d'autre ou l'intelligence se cantonne à une distribution
entre individu, comme chez les insectes sociaux. La vie peut
également
se développer uniquement au niveau microbien, avec une
grande variété d'espèces (comme sur Terre)
et quelques organismes pluricellulaires. D'autres milieux pourraient
également conduire à l'apparition d'animaux
pluricellulaires,
mais sans conduire inévitablement, sinon à
l'intelligence,
du moins à la conscience. Il en a été ainsi
sur Terre: en 200 millions d'années, les reptiles ne sont
pas parvenu à la technologie, alors qu'il a suffit de 2
millions d'année aux mammifères pour conquérir
la lune !
Pour les espèces conscientes, des phénomènes de convergence évolutives pourraient contribuer à donner à nos "frères des étoiles" un aspect proche du notre. En effet, le développement de la technologie passe par la pré-existence de caractères anatomiques voisins des nôtres: une vision du relief impliquant au moins deux yeux, des membres permettant la préhension d'objets de façon précise, la possibilité de se déplacer (sans laquelle un cerveau est inutile), une séparation entre centre nerveux et nutritifs (l'un n'entravant pas la croissance de l'autre) ect...
Tectonique des plaques
et évolution: l'influence de l'environnement
Sur notre Terre, la surface de la planète est, on ne sait pourquoi (suite à un choc ? a cause de sa teneur en eau ?) fragmentée en plaques qui se déplacent, créant en permanence de nouveaux reliefs. Cette tectonique n'est pas activement observée sur les autres planètes telluriques du système solaire (Ce qui ne veut pas dire, contrairement à une opinion trop rapide, qu'elles sont dépourvues de relief...). Elle a un rôle important sur l'évolution des espèces vivantes: maintenues isolées les unes des autres, les différentes espèces sont confrontées à un solitude reproductive qui accélère leur évolution, leur différenciation, et donc les processus par lesquels de nouvelles espèces apparaissent. Sur une planète sans tectonique, les précipitations auront tôt fait de rendre celle ci assez "lisse", sans isolement autre que la distance et les accidents créés par le réseau hydrographique.... les climats sur une telle planète seraient différents de ceux que nous connaissons, et l'évolution y serait, sans doute, plus lente que sur Terre. Bien sur, rien n'interdit non plus que la tectonique soit également à l'oeuvre sur d'autres planètes telluriques, peut être de façon transitoire (des structures géologiques laissent à penser qu'elle a été active sur Venus et Mars), avec des conséquences similaires à celles observées sur Terre...
A ce sujet, un effet étonnant, mais qui n'est peut être qu'un artefact du à notre connaissance imparfaite des fossiles: il semble que l'intégralité des plans d'organisation des animaux (mêmes ceux qui ont disparus par la suite) soit apparue à la même époque, il y a 600 millions d'années, en 50 millions d'année seulement. Que c'est il donc passé à cette époque? quelle a été l'origine de la pression évolutive qui a forcé les êtres vivants à se diversifier, se complexifier parfois, ou disparaître ?
Les chercheurs ont été conduits à incriminer une série de glaciations, des collisions avec des météores, une activité volcanique intense.... Les arguments les plus pertinents font appel à une période glaciaire extrêmement intense, maintenue pendant 10 millions d'année et ayant recouverte complètement notre planète sous un manteau de glace (Hoffman & Schrag, 2000). A cette période aurait succédé de façon très rapide une époque chaude, l'ensemble de cette alternance climatique effectuant une sélection drastique parmi les formes de vies présentent à l'époque et favorisant une diversification explosive des formes de vies.
Mais pourquoi, par la suite, une telle diversification n'a telle plus été possible ? Ce ne sont pourtant pas les extinction en masses qui ont manqué, mais sans doute n'ont elle pas atteinte l'intensité sélective de la superglaciation dite "néoprotérozoïque". Mais ceci est une autre histoire, notre histoire, dont les derniers chapitres ont été écrit sur Terre, mais dont les premiers sont partagés, à travers la voie lactée, par toutes les créatures qui peuplent notre galaxie.
roger.raynal@wanadoo.fr
Si ce travail vous a plu (ou non!),
Donnez moi votre avis
!
Des
"validations"
officielles :
Petit à petit, les idées présentées ici se répandent et sont portées à la connaissance de la communauté scientifique .
Sous l'influence de sources d'énergie variées, des molécules prébiotiques comme les acides nucléiques et les acides aminés se forment à la fois à la surface des planètes telluriques et au niveau de nombreux corps célestes (poussières, noyaux cométaires, astéroïdes). L'eau liquide ainsi que, plus lentement, la glace, favorisent la formation de polymères tels que de petits ARN et des peptides. Ces polymérisations sont catalysées par des minéraux argileux, des structures cristallines, et les ions minéraux. Cependant, les catalyseurs principaux sont représentés par les grains de poussière interstellaire qui, atteignant le milieu liquide terrestre, y permettent des synthèses plus complexes. La vie naît de la rencontre entre les acides aminés d'origine extraterrestre et les acides nucléiques majoritairement synthétisés sur Terre . Les structures moléculaires ancêtres de nos ribosomes actuels, se forment à la surface des grains de poussière interstellaire qui constituent le squelette minéral sur lequel ces structures se développent: c'est le "ribomonde".
Acides nucléiques (de type oligo-nucleotides (L)-a-threofuranosyl) et peptides évoluent conjointement à la surface de cristaux, dont les défauts, constituants la première information génétique reproductible, facilitent l'action catalitique. La croissance cristalline reproduit ces substrats réactionnels, offrant une base à la réplication des ARN et des peptides qui leur sont associés, constituant ainsi des "paléozymes" ancêtre des ribosomes actuels. Les cassures des cristaux dues aux chocs ou plus simplement l'apport continuel de poussière extraterrestre assurent la multiplication de ces paléozymes, éventuellement isolés dans des membranes protéiques ou protéo-lipidiques à base de terpènes. Les premiers ARN formés, chimiquement différents et ancêtres des ARNr et ARNt, se lient à d'autres polymères d'ARN ainsi qu'à des peptides. On obtient un "ribomonde" avec des ARNt enrobés de peptides, liés à des ARN "libres" (futurs ARNr et ARNm) et d'autres acides aminés à la surface de poussières interstellaires. Les synthèses des polymères biologiques nécessitent un milieu pauvre en eau. La sélection moléculaire s'exerce pour favoriser les ARN "libres" se réplicant rapidement et suffisamment résistants. On obtient ainsi des quasi espèces de paléoribosomes liés à des ARN. Le code génétique ne suit pas la vie: il se développe avec elle. La présence de catalyseurs cristallin permet de repliage des ARN ainsi que la réalisation de liaisons peptidiques. Ainsi se met en place un premier code génétique rudimentaire associant une base à l'un des 4 acides aminés majoritaires dans le milieu (glycine, alanine, asparagine, valine). Les processus de sélection aboutissent à une réplication généralisé des ARN. Le support cristallin perdant de son importance, celui ci disparaît (les cofacteurs enzymatiques en sont un souvenir).
On obtient ainsi les premiers cellules vivantes ressemblant vaguement aux procaryotes actuels. Il y a lors de ce processus coopération minérale/organique et coévolution des ARN et des peptides. Les bombardements météoritiques de la surface des planètes telluriques permettent la dispersion spatiale des produits de synthèse ne pouvant s'effectuer que dans ces lieux. Ceci implique une dispersion interstellaire non seulement des molécules prébiotiques mais également des premiers êtres vivant, et conduit donc à la réalisation d'une vague de dispersion et d'apparition de la vie dans une vaste région de la galaxie se produisant en un laps de temps géologiquement très court.
Ces premières formes vivantes ont pour caractéristiques une très petite taille et une forte association avec les minéraux. En cela, elles se rapprochent des micro-organismes nommés nanobes, largement répandus et récemment découverts sur Terre ainsi que dans des météorites.
Abstract:
Under the influence of varied sources of energy, prebiotic molecules as nucleic and amino acids appear at the same moment at the surface of the telluric planets and at the level of numerous celestial bodies (interstellar dusts, cometary pits, asteroids, atmosphere of giant planets). The liquid water as well as, more slowly, the ice, favor the forming of polymers such as of small RNA and peptides. This polymerisations is catalysed with clayey minerals, crystalline structures, and mineral ions.
However, the main catalysts are represented by the interstellar dust which, reaching ground liquid environment, allow more complex syntheses. Life arises from the meeting between the amino acids of extraterrestrial origin and the nucleic acids mainly synthetized on Earth. Molecular structures ancestors of our actual ribosoms, formed in the surface of interstellar dust which constitute the mineral skeleton on which these structures develop: it is the " riboworld ". Nucleic acids (of L-type oligo-nucleotides ) -a-threofuranosyl) and peptides evolve collectively at the surface of crystals, in which defects, establishing first reproducible genetic information, facilitate catalitic action . The synthesis of biologic polymers requires an hydrophobic environment.
Crystalline growth reproduces these reactionnal environment, offering a base to the replication of the RNA (or at this molecular ancestor) and the peptides which are associated to them, establishing so the " paleozyms ", ancestor of the current ribosomes. The breaks of crystals owed to shocks or more simply continual contribution of extraterrestrial dust assure the reproduction of this paleozyms, possibly isolated in mineral with proteins or terpenic membranes.
The first RNA formed, chemically different and ancestors of the actuals rRNA and tRNA, are bound at other RNA polymers as well as peptides. One obtains a " riboworld " with coated tRNA of peptides, connected to "free" RNA ( future rRNA and mRNA) and the other amino acids in the surface of interstellar dusts. Molecular selection exercises to favor the "free" ARN réplicant quickly and resistant enough. One obtains so almost paléoribosoms bound to ARN.
The genetic code does not follow life: he develops with it. The presence of crystalline catalysts allows of ARN withdrawal as well as the realization of peptidic connections. So is set up a first rudimentary genetic code associating a base to one of 4 majority amino acids in the environment (glycin, alanin, asparagin, valin). The processes of selection end in a replication generalized with the RNA. The losing of importance of crystalline support, that this disappears (enzymatic cofactors are a recollection of it).
One obtains so first alive cells looking like vaguely the modern procaryotes. There is during this mineral / organic process cooperation and coevolution of the RNA and the peptides. Meteoritic bombing of the surface of the telluric planets allow the spatial dispersal of synthetic products being able to be made only in these places. This implies an interstellar dispersal not only prebiotics molecules but also first living beings, and leads so to the realization of a wave of dispersal and appearance of the life in the galaxy occurring in a lapse of time geologically very short.
These first alive forms have for characteristics a very small size and a strong association with minerals. In it, they get closer to named micro-organisms nanobes, widely wide-spread and recently discovered on Earth as well as in meteorites.
R.Raynal