Dernière mise à jour: janvier 2006
C'est là la première question à se poser si l'on veut prendre en compte la diversité du vivant. A partir de quand pourrons-nous parler de vie? Le problème n'est pas aussi simple qu'il paraît, et classiquement on propose qu'un être vivant soit capable à la fois de naître, de se développer, de se reproduire, de se nourrir, de produire des déchets et enfin de mourir. Cet aspect des choses reste assez restrictif et, si on l'applique réellement, conduit à des impasses: une bactérie se reproduit en se divisant, elle ne meurt pas ainsi: immortelle, elle ne serait donc pas vivante! De même, si l'on considère que la division cellulaire est une forme de mort, alors les cellules ne se reproduisent pas, et ne sont pas vivantes !
Même imparfait, cet ensemble de conditions permet d'éliminer certains processus qui miment la vie sans pour autant, si l'on s'en tient là, en faire partie. Ainsi, les cristaux qui apparaissent dans une solution saturée sont-ils capables de croître, se nourrissant des éléments de leur milieu ambiant, de se multiplier éventuellement, et de "mourir" s'ils sont dissous. Le statut des virus pose aussi problème: ils ne sont pas capables de se reproduire, mais on ne peut leur dénier une parenté plus que certaine avec les autres formes de vie. Il faut bien voir que l'environnement possède une importance énorme dans la définition de la vie: une fois dans une cellule, un virus peut être considéré comme vivant, et non en dehors. Telle molécule capable de se reproduire en tube à essai se révélera complètement inerte en milieu naturel etc... Cette importance de l'environnement peut être illustrée par l'exemple des parasites d'ordinateurs que sont les virus informatiques. Si on considère un environnement purement informatique, cybernétique, dont la manifestation physique n'est que la modification de l'orientation magnétique d'un fragment de disque dur ou l'état électrique d'un composant de mémoire, alors nous pouvons valablement soutenir que les quelques lignes de code qui constituent un virus informatique sont bien près, dans ce "biotope" particulier, de la vie. Il en est de même pour les "agents", ces programmes qui sont envoyés sur le réseau internet afin d'y trouver et d'y sélectionner les informations utilisables par les moteurs de recherche.
Il me semble qu'une autre définition de la vie, plus adaptée à la description de ses manifestations primitives, peut être utilisée: on considère comme vivant tout système suffisamment organisé pour être en interaction avec son environnement, se perpétuant dans le temps en se reproduisant de façon imparfaite, de manière à pouvoir évoluer, et qui est soumis à des pressions sélectives de la part de son milieu.
Une autre définition, plus classique, en a été donnée par MD Morch: " est vivante toute structure capable de s'organiser, d'assimiler et de transformer des éléments simples en macromolécules complexes, et capable de se reproduire à l'identique à elle-même." Cette façon de voir est assez restrictive puisqu'elle ne prend pas en compte l'environnement, et suppose une reproduction à l'identique qui ne reste q'une vue de l'esprit...
La vie semble bien liée inéluctablement à des phénomènes de reproduction imparfaite (ce qui nous permet d'en exclure les virus informatiques préalablement décrits). Elle va donc prendre sa source dès qu'une structure, une molécule unique, un complexe plurimoléculaire ou une chaîne de réactions chimiques, peu importe, seront capables de se perpétuer de façon imparfaite.
Les êtres vivants aujourd'hui
L'étude de la biochimie des êtres vivants nous a confirmé, ces 50 dernières années, ce qu'avait permis de supposer l'étude des ressemblances morphologiques et anatomiques des différents groupes, ainsi que les indications de la paléontologie: toutes les formes vivantes possèdent une biochimie comparable, basée sur des molécules similaires et sur un unique système de transmission de l'information d'une génération à une autre. Cela implique que toutes ces formes de vie dérivent d'un ensemble homogène d'organismes primitifs, peut-être un seul, peut-être plusieurs. Les biologistes qui ont classé les êtres vivants, ce qui est très difficile à réaliser, ont cru bon parfois de se dire que leur classification allait des êtres les plus petits, les plus "simples" qu'ils disaient moins évolués, vers les plus évolués, à savoir bien sûr les mammifères et, tout en haut, l'être humain. Cette conception, que l'on retrouve encore dans nombre d'ouvrages de vulgarisation, est d'un anthropocentrisme confondant: contrairement à ce que l'on pourrait penser, la méduse n'est pas "moins évoluée" que l'être humain, bien au contraire: les différentes espèces de cet animal gluant ont réussi à traverser des crises majeures et des cataclysmes de l'histoire de la Terre qui aurait suffit à effacer de sa surface tout le groupe des mammifères! Si nous voulons nous interroger valablement sur la vie, alors il ne faut pas nous hausser sur un piédestal improbable et regarder de haut les limaces, les mousses et les bactéries: l'aune de l'évolution se mesure à l'horloge du temps passé, et au point de vue évolutif la moindre des bactéries qui grouille dans vos intestin dispose de 3 milliards d'années d'avance sur les premiers êtres vivants pluricellulaires!
Gardons cela à l'esprit et voyons donc quel sont les différents groupes dans lesquels on peut ranger, de façon très générale, les êtres vivants:
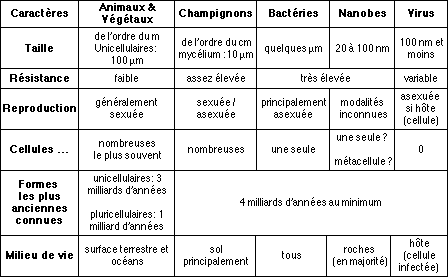
Ces trois groupes font partie de ce que l'on appelle les eucaryotes: leurs cellules sont grandes, compliquées, et contiennent tout un ensemble de compartiments internes. Les trois autres groupes, constituant les procaryotes, sont beaucoup plus simples, plus évolués et bien plus résistants. Ils différent beaucoup des eucaryotes, à un point tel qu'il est même possible qu'ils ne possèdent pas la même origine, ce qui s'explique, nous le verrons, dans notre scénario sur la genèse de la vie.
| Organismes | bactéries | archaebactéries |
| Génome Transposons (éléments génétiques mobiles) Introns (ARN non codant) ARN polymérases Paroi cellulaire Acides gras Lipides |
stable rares non simples peptidoglycanes linéaires liaison ester, contiennent du L-glycérophosphate |
variable
(réarrangements fréquents) nombreux oui (ARNr et de transfert) grandes, avec sous unités, ressembmant à celles des eucaryotes pas de peptidoglycane ramifiés liaison éther, contiennent du D-glycérophosphate Ribosomes, tRNA et r RNA avec caractères mixtes eucaryotes/bactériens. |
|
Les
nanobes ont tout d'abord été identifiés dans des
carbonates précipitant au niveau de sources thermales chaudes
par Robert L. Folk, professeur émérite de géologie
à l'université du Texas, en 1992. Observés sous la
forme de filaments ou de sphéroïdes, leur nature biologique
est alors seulement suspectée. Plusieurs chercheurs* identifient
ensuite des structures semblables, interprétées comme
étant les fossiles de formes de vie primitives, toujours
associées fortement à des minéraux tels que les
argiles, les sulfites, les silicates (que l'on retrouve dans la
poussière interstellaire).
Ces "structures biomimétiques" sont impliquées dans les processus de minéralisation, que l'on avait toujours cru jusqu'ici être de nature abiotique. Elles ont été également identifiées dans le sang, au niveau des calculs rénaux, des plaques athéromateuses et, probablement, de la plaque dentaire. Cependant, leur étude et leur identification difficile dans les pathologies humaines sont à l'origine d'une polémique. La raison n'en est pas uniquement scientifique, mais met en jeu, comme souvent, des querelles de clochers: les nanobes, identifiés au départ sur des critères purement morphologiques, voient leur statut biologique nié par des chercheurs les considérant comme des artefacts ou des formations d'origine strictement minérale. Le découvreur des nanobes, le Pr Folk, signale que ceux-ci sont tellement nombreux dans certains échantillons de roches qu'il est fort possible que la biomasse des nanobes surpasse celle de toutes les espèces vivantes à la surface de la Terre. En effet, ces "organismes" présentent la caractéristique de pouvoir résister à de très fortes pressions et températures. Le statut biologique des nanobes est cependant établi en 1998 par une étude parue dans la revue American mineralogist et réalisée par le Pr. Philippa J.R. Uwins et son équipe, de la Queensland university, basée sur des nanobes provenant de grès recueillis lors de forages pétroliers off shore à plus de 4000 m sous la surface océanique, soit à des températures d'environ 150°C et des pressions de 2000 atmosphères!. Les résultats obtenus montrent:
J'ai pris contact avec le Pr. Uwins qui a eu l'amabilité de me tenir au courant de ses recherches actuelles, en voie de publication, selon lesquelles la nature biologique des nanobes ne fait plus aucun doute. Bien que la préparation de ces organismes très résistant et hydrophobe pour la microscopie électronique à transmission soit très ardue, l'équipe de ce chercheur a obtenu, en améliorant ses techniques, les résultats suivants:
Il s'agit donc bien de micro-organismes inconnus jusqu'à ce jour... Le milieu scientifique français est, à propos de cette découverte, d'un silence éloquent... La seule trace que j'ai trouvée est un entrefilet d'une quinzaine de lignes dans le n° 251 de la revue Pour la Science, concernant la possible origine infectieuse des calculs rénaux, et un bref article dans Science & vie de janvier 2000. Pourquoi ce manque d'enthousiasme? Tout simplement parce que les nanobes sont trop petits pour contenir la machinerie moléculaire assurant la vie des bactéries. Leur biochimie doit donc être considérablement différente de celle des autres organismes que nous connaissons... Un autre élément explique peut-être ce silence gêné: les nanobes ressemblent beaucoup aux structures découvertes dans plusieurs météorites, dont nous reparlerons, et qui ont été considérées comme non probantes à cause, en particulier, de leur taille insuffisante... On est en présence d'un faisceau d'indices convergents visant à établir l'existence de formes de vie primitives, liées aux minéraux, et répandues largement dans tout l'univers. Nous verrons que ceci cadre parfaitement avec mes conceptions donnant à la vie sur notre planète une origine essentiellement extra-terrestre avec une forte implication des minéraux. |
"Notre"
monde est surtout celui des micro-organismes
Notre taille et les limites de notre vision nous ont conduit à ne voir pendant des millénaires à la surface de la Terre que des organismes pluricellulaires. L'invention et le perfectionnement du microscope optique ont permis de découvrir le foisonnement des micro-organismes, en particulier des bactéries, dont l'implication dans les pathologies infectieuses a été un puissant stimulant de la recherche. La mise au point du microscope électronique a permis de découvrir les virus, puis les dernières évolutions du microscope électronique à balayage ont montré ces dernières années l'existence des nanobes. Au fur et à mesure que nos techniques d'observation progressaient, l'importance de l'être humain, puis des mammifères, puis des êtres pluricellulaires décroissait. Si l'on s'intéresse au nombre d'espèces, d'individus ou à leur dispersion dans tous les milieux, les micro-organismes constituent l'essentiel de la vie sur Terre. Ce sont eux qui ont fourni de l'énergie aux ancêtres des cellules eucaryotes, eux qui ont commencé à produire l'oxygène de notre atmosphère, eux qui permettent aux animaux de dégrader les molécules des végétaux, eux qui fabriquent, dans notre intestin, certaines molécules qui nous sont indispensables. Outre les utilisations industrielles que nous en faisons, les micro-organismes sont les représentants majeurs de la vie sur Terre, et ils ont "pris le départ" il y a environ 4 milliards d'années au bas mot, évoluant et se perfectionnant seuls pendant 3 milliards d'années... Quand on réalise qu'une durée 6 fois moindre a suffit pour passer des premières éponges à l'homme, on mesure le chemin évolutif parcouru par les ancêtres des bactéries, et combien il nous est difficile d'imaginer leur ancêtre: celui-ci est sans doute plus différent d'une bactérie actuelle qu'une éponge ne l'est d'un être humain!
Des tentatives de dénombrement (imprécises par nature) du nombre de cellules de micro-organismes, excluant les nanobes, donnent le chiffre fantastique de 5x1030 unités bactériennes sur notre planète, dont plus de 90 % résident dans le sous-sol des océans ou des continents! Ce nombre fantastique pose en lui-même un problème, car la quantité de carbone disponible ne permettrait pas le renouvellement de ces populations bactériennes à la vitesse à laquelle on l'observe. Les Pr. Whitman et Wiebe, de l'université de Géorgie, aux USA, en déduisent que les bactéries profondes doivent être capables d'utiliser des sources d'énergie jusqu'ici inconnues. Confortant cette hypothèse, l'équipe de F. Chapelle, de l'US geological survey, a identifiée en janvier 2002 une communauté de micro-organismes vivants 200 m sous le sol de l'Idaho, dans des sources chaudes, et tirant leur énergie de l'hydrogène contenu dans les roches (ce qui les conduit à rejeter du... méthane!).
On lit souvent dans les ouvrages de vulgarisation, mais aussi dans nombre de traités s'adressant à un public plus restreint, que l'évolution posséderait un "sens", allant toujours vers l'édification de structures de plus en plus complexes. Ce dogme de la complexité croissante se retrouve également dans d'autres domaines, en astrophysique par exemple. Cependant, pour ce qui est des êtres vivants, il est erroné, c'est une illusion d'optique due à la valorisation indue que nous accordons aux formes de vie qui nous ressemblent. Si l'on regarde l'évolution temporelle des formes de vie sur Terre, on constate qu'a partir d'un certain niveau d'organisation indispensable pour que l'on parle de matière vivante (et que l'évolutionniste Stephen J Gould, paléontologue à l'université d'Harvard, appelle le "mur de la complexité"), le développement de formes de vie complexes, pluricellulaires, ne constitue pas une règle mais plutôt une exception: ce qui évolue, c'est surtout le nombre, l'adaptation et la répartition des micro-organismes, les métazoaires n'en constituant qu'un diverticule certes intéressant pour nous, qui sommes concernés au premier chef, mais peu significatif au point de vue de la diversité des formes vivantes. De plus, les métazoaires restent confinés à des milieux bien spécifiques, car ils sont plus fragiles, et leur évolution est plus lente, que celle des procaryotes.
Le cas des virus mérite un examen spécifique: on considère actuellement que ces "êtres" constituent la forme la plus achevée de l'évolution des procaryotes, ceux-ci se limitant alors à la simple perpétuation d'une information génétique au dépend d'autres cellules. Le fait que les virus soit incapables de se reproduire seuls a fait croire qu'ils ne pouvaient pas être les ancêtres des autres êtres vivants, mais rien n'interdit de penser qu'ils pourraient provenir qu'une "involution" secondaire, leur mode de vie parasitaire, leur ayant fait perdre des caractéristiques ancestrales assurant leur reproduction. Des "régressions" semblables s'observent couramment chez les animaux menant une vie parasitaire, et dont il ne subsiste parfois qu'un simple organe reproducteur. Bactéries et archaebactéries peuvent être infestées par des virus, ce qui témoigne de leur ancienneté. L'étude des gènes viraux révèle également que ces derniers sont non seulement très anciens mais aussi qu'ils ont pu jouer un rôle majeur dans l'évolution des organismes, allant même jusqu'à remplacer des gènes bactériens ou eucaryotes, à moins qu'ils n'en dérivent: les rapports entre les virus anciens et les premières formes de vie sont complexes, mais indubitablement réels (une grande partie de l'ADN "muet" de nos propres cellules est très probablement d'origine virale).
De nombreux micro-organismes supportent des conditions de vie d'une exceptionelle dureté

Les procaryotes
ne connaissent pas la reproduction sexuée au sens où
nous l'entendons: chez eux, il n'y a pas brassage de gènes
et recomposition d'un nouveau génome mais échange
direct de fragments de
matériel génétique. Ajouté à
leur fantastique capacité de prolifération qui
accélère
encore leur évolution, cet échange
génétique
leur a permis de s'adapter à strictement tous les milieux
que nous avons pu découvrir sur notre planète, et
de résister à un séjour de quelques années
sur la Lune. Ainsi, 2 chercheurs de la Nasa (R. Hoover et E. Pikuta)
ont découvert une nouvelle espèce
bactérienne
nommée Tindallia
californiensis , au niveau
du lac Mono (très salé et basique, ùais abritant
de nombreux organismes), en Californie. Cette bactérie
prolifère dans un milieu comprenant 20 % de sel et à pH
10 !
Si nous voulons savoir comment sont apparus tous ces êtres
vivants, à commencer par les plus simples, les bactéries,
il va falloir nous intéresser à leurs
éléments
constitutifs, les molécules du vivant. Nous verrons que
certaines d'entre elles correspondent à celles que nous
avons déjà rencontrées dans le milieu
interstellaire,
et que cette correspondance est un indice supplémentaire
de l'origine extraterrestre de la vie.
Des polymères, une structure
Les plus importantes de ces molécules sont de très grandes tailles: on parle de macromolécules. Elles sont formées de l'enchaînement de molécules plus petites, les monomères. En simplifiant beaucoup, on peut distinguer 3 grands groupes de macromolécules caractérisant le vivant:
Les polysaccharides sont formés de l'enchaînement de plusieurs centaines à plusieurs milliers de sucres (le plus souvent, du glucose). Ils constituent généralement des réserves énergétiques alimentaires (comme l'amidon des végétaux, ou le glycogène de notre foie) ou des protections vis-à-vis du milieu extérieur (cellulose des végétaux, chitine de la carapace des insectes et crustacés). La façon dont les chaînes de sucres sont "branchées" les unes sur les autres conditionne les propriétés des macromolécules obtenues. Certaines de ces macromolécules sont parmi les plus abondantes sur Terre (cellulose).
Les protéines sont des enchaînements d'acides aminés et constituent l'essentiel des structures non informatives des cellules. Bien qu'il existe plusieurs dizaines d'acides aminés différents, les êtres vivants n'en utilisent que 20 (hormis quelques rares micro-organismes à la paroi spécifique). La forme d'une protéine conditionne sa fonction: la longue chaîne des acides aminés se replie dans l'espace, ces derniers pouvant se lier les uns aux autres. Le plus souvent, la chaîne d'acides aminés dessine dans l'espace une hélice ou une suite de plans appelés feuillets. Ces structures se replient encore pour donner sa forme à la protéine. Ces molécules peuvent jouer des rôles très différents: squelette interne de la cellule, enzymes dont la forme permet de rapprocher des réactifs et accélère les réactions chimiques, signaux pour les autres cellules, récepteurs capables de capter les messages moléculaires envoyés par d'autres cellules...
La fabrication des protéines nécessite l'exploitation d'une information contenue dans d'autres macro-molécules, les acides nucléiques.
L'ADN et l'ARN contiennent et transmettent les informations qui permettent la reproduction et le fonctionnement des cellules. Ils sont formés de l'enchaînement d'acides nucléiques. Il existe 5 acides nucléiques utilisés par les êtres vivants. L'ARN est capable de se répliquer lui-même et fut sans doute le premier acide nucléique. Pour le fabriquer, il faut disposer d'un sucre, le ribose, de phosphate et de 4 bases différentes. La morphologie de ces molécules est extrêmement importante et explique leur importance. L'ADN est une molécule "double", constituée d'une hélice liée à son image, sa copie: c'est donc une hélice double, ce qui lui permet de se multiplier. Cette double hélice est entortillée plusieurs fois sur elle-même, comme un cordon de téléphone ou une corde à noeuds, et se lie à des protéines. Sa forme change continuellement dans l'organisme, ce qui complique son étude! L'ARN est lui constitué d'une seule file d'acides nucléiques qui se replient sur eux-mêmes, tout comme les acides aminés d'une protéine. Les ARN possèdent donc une forme spécifique, et sont de longueur très variable: il existe plusieurs types d'ARN, selon leur fonction.
Il existe un lien très fort et très ancien entre les protéines et les acides nucléiques. Au plan moléculaire, on peut dire que ces deux espèces vivent en symbiose, chacune étant indispensable à la survie et à la perpétuation de l'autre. Le mécanisme de la synthèse des protéines est complexe, mais nous pouvons en donner les très grandes lignes car, conservé chez tous les êtres vivants connus, il nous fournit un "fossile" supplémentaire pour l'origine de la vie. Voyons donc comment les choses se passent de nos jours, et ce que cela impliquait à l'origine:
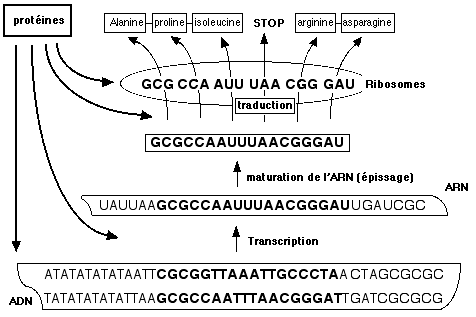
L'enchaînement des acides nucléiques dans l'ADN forme un code qui permet de faire correspondre à une suite de 3 acides nucléiques un acide aminé. Le déroulement de l'hélice d'ADN et sa lecture nécessitent de nombreuses protéines. Parmi ces dernières, certaines (les ARN polymérases) effectue une copie partielle de l'ADN en un ARN dit messager (ARNm). Le fragment d'ADN qui a été lu correspond le plus souvent à un gène. Ce gène va permettre la production d'une protéine, si celle-ci est nécessaire. L'ARNm contient de vastes régions sans signification, que l'on appelle des introns. Il se replie sur lui-même, découpe ces zones non codantes et recolle les extrémités. Ce procédé, typique des cellules eucaryotes, est appelé épissage. Ensuite, l'ARNm va être lu par d'autres ARN liés à des protéines dans des organites appelés ribosomes. Au niveau de ce complexe multi-moléculaire, des ARN ribosomiaux, dont nous reparlerons, vont permettre d'associer à chaque triplet de base un autre ARN, dit de transfert (ARNt) lié à un acide aminé particulier. La lecture à la chaîne des triplets de l'ARN aboutit donc à la synthèse par les ribosomes d'une chaîne d'acides aminés, autrement dit une protéine. Un seul ARNm peut être lu plusieurs fois, par plusieurs ribosomes en même temps: ceci permet d'amplifier le message émis par l'ADN.
Le rôle de l'ADN a donné lieu à de nombreux fantasmes, et beaucoup d'erreurs courent à son sujet dans de nombreux livres de vulgarisation. Il faut bien voir que cette molécule ne se présente pas, comme on le dit souvent, comme un livre qu'il suffit de lire et de décoder pour comprendre: l'ADN des animaux comme l'homme est en grande partie... inutile! Il ne contient qu'à peine 10% de séquences codantes pour des protéines, perdues dans un fatras d'acides nucléiques dont nous ignorons le rôle et l'importance. Certaines séquences sont probablement impliquées dans le contrôle de la forme de la molécule, dont nous verrons l'intérêt.
Plutôt qu'un livre, il faudrait imaginer l'ADN eucaryote comme un annuaire téléphonique japonais dans lequel on aurait collé, à des endroits choisis au hasard, quelques fragments d'un texte en français... Si l'on veut faire une analogie informatique, je dirais que l'ADN est un bien mauvais disque dur, où l'information est toute fragmentée en blocs multiples et éloignés, séparés par des zones sans importance (du moins en apparence). Pour pouvoir être traduite, cette molécule est contrainte à des contorsions acrobatiques, permettant de "neutraliser" les séquences non codantes. Les bactéries possèdent un ADN bien plus évolué, débarrassé de toutes les séquences polluantes et entièrement efficace: souvent, il n'a pas fini d'être transcrit en ARNm que ce dernier est déjà lu par les ribosomes et traduit en protéines!
Circulation de l'information dans les cellules
Un des dogmes de la biologie moléculaire est que l'information dans les cellules circule à sens unique: de l'ADN vers l'ARN puis vers les protéines. La découverte des rétrovirus a porté un premier coup à cette conception. On avait montré que de l'ARN pouvait donner de l'ADN, ce qui a ouvert une immense voie de recherche et d'expérimentation à la biologie moléculaire. Par contre, imaginer une communication, un transfert d'information, entre protéine et ADN ouvrirait la porte aux vieilles conceptions de l'hérédité des caractères acquis, et est ressenti comme une hérésie, conférant un statut ambigu à celui qui oserait s'en prévaloir pour expliquer des phénomènes résistant à l'analyse classique. Un exemple nous en est donné par la découverte des protéines infectieuses, les prions, par le biochimiste S. Prusiner: le fait qu'une protéine puisse transmettre une information (sa forme), se reproduire en quelque sorte et déclencher ainsi une maladie a été accepté à reculons, beaucoup de protestations ayant accueilli l'attribution du prix Nobel au Pr Prusiner, et certains, particulièrement au CEA, en France, où notre ouverture d'esprit est proverbiale, tiennent encore absolument à incriminer un virus aussi classique que discret. Ce dahu biochimique porte même un nom: le virino.
Et pourtant, si un passage d'information direct entre protéine et ARN, une "traduction inverse" ne semble pas (ou plus, la nuance est d'importance!) possible, il est indéniable qu'à chaque seconde un courant d'information circule des protéines vers l'ADN: nombre de protéines se lie à cette molécule et contrôle son activité en fonction des informations reçues à la fois de l'intérieur mais aussi de l'extérieur de la cellule. La seule concession au dogme en vigueur est que les différentes protéines de contrôle de l'expression des gènes ne modifient pas la séquence de l'ADN en réponse aux variations du milieu, empêchant ainsi qu'un caractère acquis ne devienne héréditaire. Cependant même cette règle n'est pas strictement respectée: les enzymes qui assurent la réparation de l'ADN lors de sa réplication ne sont pas infaillibles, et qu'est-ce qu'une mutation sinon, justement, un caractère acquis transmis de façon héréditaire?
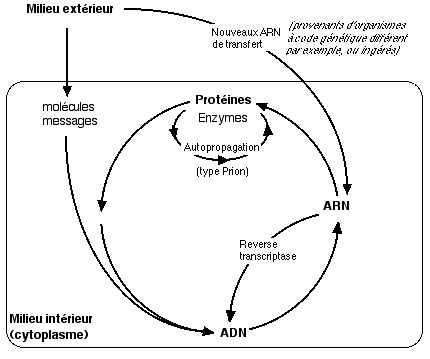
Circulation de l'information
dans la cellule: bien
que l'ADN soit le dépositaire de l'information
génétique,
celle-ci circule continuellement dans la cellule et est
influencée
par des molécules en provenance du milieu extérieur.
Si ces molécules sont des ARN de transfert, l'information
peut "remonter le courant" des acides aminés
vers l'ARN, puis l'ADN.
Il faut bien voir que les mutations se produisent apparemment au hasard, ce qui limite leur portée adaptative: la cellule ne réagit pas à un environnement donné, mais à une situation. Cet effet de l'environnement cellulaire sur le matériel génétique apparaît pleinement dans les techniques de clonage: lorsque l'on intègre le noyau d'une cellule de peau par exemple dans le cytoplasme d'un ovocyte, c'est ce dernier qui va agir sur l'ADN pour le "reprogrammer" et permettre l'expression des gènes qui vont conduire à la formation d'un embryon en lieu et place des gènes activés dans la cellule d'origine du noyau. Dans ce cas, l'ADN se comporte moins comme un programme que comme une banque de données. La lecture de ces données peut d'ailleurs se faire au hasard, sans déterminisme excessif: le Dr Jean Jacques Kupiec, du laboratoire de génétique et de virologie moléculaire de l'hopital Cochin, pense que les molécules qui contrôlent l'activité des gènes pendant le développement se déplacent au hasard le long de la molécule d'ADN, comme une perle glissant en avant ou en arrière sur un fil. Ce processus serait à l'origine de la différenciation des cellules en types distincts (cellule de peau, de foie ect...) qui seraient maintenue par des informations stabilisatrices échangées entre cellules. Cette conception ouvre la voie à la possibilité de transformer chez l'adulte un type de cellule en un autre. Cette transformation, qui aurait fait hurler de rire un jury de thèse il y a dix ans, est maintenant activement recherchée, obtenue et étudiée à cause de ses implications thérapeutiques potentielles.
Un autre exemple de la plasticité de l'information génétique nous est fourni par des gènes HOM qui guident le développement embryonnaire. Il se trouve que ces gènes sont similaires entre des organismes très éloignés évolutivement, comme la mouche et la souris par exemple. Simplement, les vertébrés ont 4 copies de ces gènes, et les mouches, une seule. C'est ce que l'on appelle une "orthologie" (homologie génétique dans des espèces très différentes). Là ou cela devient intéressant, c'est que les gènes de la souris peuvent parfaitement remplacer ceux de la mouche, et conduisent au développement du système nerveux... d'une mouche!
Les gènes qui régulent le développement du cerveau sont encore plus étonnants: nommés otd chez la mouche, ils existent chez la souris sous forme de plusieurs copies, les otx. Imaginez que l'on enlève les gènes otd à un embryon de mouche: tout l'avant du système nerveux ne se développe pas. Imaginez ensuite que l'on remplace les gènes otd (mouche) par les otx (souris): ceci a été fait, et on obtient alors... des mouches normales! Autrement dit, le même ADN qui chez la souris guide la formation d'un cerveau de souris va, dans une cellule de mouche, guider la fabrication d'un cerveau de mouche... Ceci nous montre, outre l'identité des molécules qui guident l'expression des gènes chez ces deux espèces, que l'expression de l'information contenue dans l'ADN est totalement dépendante du milieu dans lequel se trouve cette molécule.
L'ADN n'exprime donc que des potentialités qui ne prennent leur sens que dans un milieu particulier, lequel communique avec le génome via des intermédiaires que sont les molécules messagers qui circulent dans les cellules. Il n'existe pas de gène nu! (*) Une telle communication se retrouve chez toutes les formes vivantes, ce qui nous indique que dès le début de la vie acides nucléiques et protéines n'ont pu qu'évoluer de concert. Nous verrons que la plus ancienne structure à avoir conservé les traces de cette évolution est le ribosome, qui selon nous est le reliquat d'une des toutes premières formes de vie. Nous pourrons avancer alors une hypothèse permettant d'expliquer pourquoi les acides nucléiques se sont mis à coder pour des acides aminés, ce que personne jusqu'ici n'a été en mesure d'expliciter.
(*) : Toutefois, d'étonnantes formes de "vie"(?), les viroïdes, sont de bons candidats au titre de "gènes nus" actuels. Ce sont des anneaux d'ARN, simple brin, ne contennant que quelques centaines de nucléotides. Il sont capables de pénétrer dans les cellules végétales et de s'y reproduire. Leur nom fait souvent référence à la plante qu'ils attaquent (potato spindle tuber viroid) où à la malide qu'ils provoquent (cadang-cadang disease viroid). Cependant, les viroïdes ne codent aucune protéine (ce ne sont donc pas vraiment des gènes!) mais se contentent de se réplicer eux même ad infinitum...
Les molécules du vivant tirent leur spécificité de leur forme, mais aussi des mouvements que ces dernières autorisent ou interdisent, ils sont donc dépendants de leur topologie. Cette "dictature de la forme" a été expérimentalement vérifiée chez les protéines, dont le repliement est plus affaire de topologie que de propriétés chimiques. Des molécules de séquences différentes mais possédant des formes similaires peuvent donc posséder des activités identiques.
Pour qu'une molécule soit capable de s'auto-répliquer, sa forme est primordiale ainsi que la répartition des forces exercées par les différents groupements moléculaires qui la composent. Ces forces peuvent orienter et attirer les molécules de base, et la forme de la molécule permettre le rapprochement des diverses molécules afin de faciliter leur réaction: c'est le mode de fonctionnement des enzymes de nos cellules. Pour ce faire, l'attitude expérimentale qui consiste à essayer d'obtenir des molécules de synthèse capable d'auto-replication à montré la prééminence des acides nucléiques: les molécules de synthèse qui peuvent catalyser leur reproduction, évoluer et se disputer les ressources de leur milieu contiennent des acides nucléiques. C'est le cas de l'ARNI (adénine ribose naphtalène imide) qui peut se répliquer ou s'associer avec elle-même, ou de la diaminotriazine xanthène thymine (DIXT) qui peut se lier et engendrer par recombinaison d'autres molécules douées d'autoreplication ou non. Cela montre l'intérêt de l'adénine, molécule présente dans les deux composés précédents et dont les mécanismes de synthèse sont bien connus dans différents milieux abiotiques.
De même, les activités de l'ADN sont largement tributaire de la forme que peut adopter cette molécule: les nuds et les entortillements qu'elle forme permettent d'exposer ou de dissimuler des régions codantes, ce qui participe à la régulation de l'expression des gènes. Il existe en effet plusieurs variantes différentes de la double hélice d'ADN. Si la plus connue est la forme dite B, il en est une autre, la forme Z, où la direction de l'axe de la molécule est modifiée. On a pu montrer que des protéines sont capables d'effectuer la transition entre ces deux topologies de la molécule. Expérimentalement, on peut aller bien plus loin encore dans la déformation de l'ADN: en provoquant la torsion de brins de cette molécule, des chercheurs de l'école normale supérieure de Paris sont même arrivés à un véritable "retournement de l'ADN", les bases normalement enfouies au centre de la molécule se retrouvant exposées à l'extérieur de celle-ci.
Le fait que les acides nucléiques forment des "rubans" capables de se replier sur eux-mêmes à cause des liaisons de nature électrique entre les acides nucléiques, en adoptant une forme particulière, explique que si leur morphologie dépend en partie de leur séquence, leur activité dépend toujours de leur forme. Il en est de même pour les protéines, dont la forme est également capable de transférer une information. La seule différence semble être que l'information transmise par la séquence des acides nucléiques est pérennante alors que celle provenant des protéines est labile, et se limite au milieu interne de l'individu, sans se transmettre, le plus souvent, aux individus de la génération suivante.
Un étrange liquide bien mal connu: l'eau
Si l'on excepte les molécules situées dans la membrane lipidique des cellules, la plupart des polymères biologiques sont en solution dans un liquide commun mais étrange, l'eau. En effet, si l'on se réfère à sa formule chimique, l'eau devrait bouillir autour de 0°C et ne se solidifier qu'autour de -100°C. Pourquoi se comporte-t'elle autrement? Chaque molécule d'eau a tendance à s'accrocher faiblement par tous les moyens à ses voisines. A température ambiante (20°C), elle change plusieurs milliers de fois de partenaires par seconde mais l'ensemble des forces générées par toutes les molécules qui frétillent ensemble permet à ces dernières de rester jointives: l'eau reste liquide à température ordinaire. Cette agitation moléculaire est si complexe qu'à l'heure actuelle la structure précise de l'eau à l'état liquide est inconnue, ce qui peut sembler étonnant; la complexité peut se cacher parmi les substances les plus courantes!
Tout comme les molécules d'eau ont tendance à s'accrocher entre elles, elles se lient aussi de façon transitoire à toutes les molécules solubles: un ion sodium par exemple se déplace entouré d'une véritable armure de molécules d'eau qui se relaient en permanence à son voisinage, formant ce que l'on appelle une couche de solvatation qui augmente son diamètre. Cette armure d'eau n'est pas sans conséquence sur les processus biologiques: une protéine contient de nombreux groupements chimiques, plus ou moins chargés, et se comporte dans l'eau comme un champ de force mobile dont certaines régions attirent les molécules d'eau alors que d'autres ont tendance à les repousser. La protéine se tortille, les molécules s'agitent et tout cet ensemble échange en permanence de l'énergie sous forme électromagnétique. Il en résulte que la présence de l'eau n'est pas neutre pour les molécules du vivant: elle conditionne leur forme mais joue aussi un rôle encore mal connu dans leurs échanges mutuels. Les molécules d'eau peuvent ainsi diminuer ou augmenter le "relief" d'une région moléculaire, plusieurs milliers de fois par seconde, et les relations intermoléculaires se font toujours à travers cette armure d'eau dont on commence tout juste à soupçonner l'importance.
La glace d'eau possède aussi des propriétés intéressantes: les molécules d'eau y sont plus ou moins figées selon une douzaine configurations possibles, ce qui permet une grande variété de comportement de ce solide et assure des conditions variées pour les molécules qu'il peut emprisonner. Ces dernières, entre autres, pourront ainsi conserver dans le milieu interstellaire une certaine mobilité, ce qui facilite parfois les réactions chimiques à basse température. Ce fait a été vérifié par S. Miller, qui a obtenu a partir d'une solution de cyanure d'ammonium oubliée 27 ans dans un congélateur à -80°C des bases puriques et pyrimidiques entrant dans la composition de l'ARN.
Un métabolisme, une dynamique énergétique
Bien que cette façon de voir soit assez peu enseignée, un être vivant ne peut se limiter à une collection de molécules et de mécanismes: c'est avant tout, au plan physique, un convertisseur d'énergie en polymères. L'origine de cette énergie est extrêmement variée, mais elle sert, en définitive, à une chose: incorporer dans les molécules des cellules du carbone provenant de l'environnement. Les voies d'utilisation et de conversion de cette énergie constituent le métabolisme, dont l'étude sort de notre domaine: retenons qu'il s'agit d'une suite de réactions chimiques permettant de fabriquer de nouvelles molécules, de dégrader les anciennes et d'obtenir l'énergie nécessaire à ces deux occupations. Il existe peu de voies métaboliques par rapport à la diversité des êtres vivants, ce qui nous donne une nouvelle indication sur les débuts de la vie. On peut donc à partir de l'étude des métabolismes actuels tenter de remonter le temps vers le premier métabolisme! Le tout est ce savoir si les premiers êtres vivants étaient autotrophes ou hétérotrophes. Le chimiste G. Wachtershauser, faisant l'hypothèse de l'autotrophie, aboutit à un métabolisme primitif ou le carbone est fixé dans les organismes en utilisant des composés minéraux soufrés, comme le sulfure de fer. Le premier être vivant serait donc, comme l'appelle ce chercheur, un "métaboliste" confiné dans une structure minérale à deux dimensions, celle des cristaux. Cette conception d'une vie primitive bidimensionnelle est extrêmement intéressante, car elle évite la "dilution" fatale aux premiers organismes "libres" dans l'eau.
Il n'est cependant pas absurde de faire le pari de l'hétérotrophie de certaines formes de vie primitives: la matière organique d'origine interstellaire et terrestre étant abondante à l'époque, certains êtres vivants auraient pu s'en nourrir, faisant ainsi l'économie d'un mécanisme biochimique complexe d'assimilation du carbone. Ce n'est que lorsque les molécules organiques se seraient faites plus rares que la pression de sélection aurait favorisé les autotrophes et les hétérotrophes capables de se nourrir de ces derniers. Les hétérotrophes ancestraux, ne trouvant plus à se nourrir, se seraient alors réduits à l'état de spores, qui au fil de l'évolution auraient pu aboutir aux plus simples et aux plus efficaces des édifices moléculaires du "paravivant": les virus.
Un organisme vivant ne peut, du fait des lois de la physique, prétendre à l'immortalité totale. Bien que les phénomènes de la sénescence et du vieillissement soit extrêmement peu connus, les unicellulaires ont trouvé un moyen très simple de perdurer: un individu recopie son génome, puis se divise pour en donner 2 (ou une copie de lui-même, c'est selon le point de vue adopté!). Au plan moléculaire, cela implique une transmission d'une information génétique par réplication. Une molécule est capable de transmettre une information, en fait, si elle est capable de reproduire sa forme. Sur le support de cette information génétique, le débat, mine de rien, reste ouvert:
· On peut émettre l'hypothèse que, comme de nos jours, ce sont les acides nucléiques qui ont toujours servi de base à l'information génétique. Longtemps, les chercheurs se sont demandés qui des enzymes ou des acides nucléiques étaient apparus en premier: nous avons vu que ces deux familles de molécules semblent indispensables l'une à l'autre. Ce petit jeu de la poule et de l'oeuf dura jusqu'en 1986 où le biochimiste T. Cech observa chez un protozoaire que l'ARN peut être son propre enzyme, catalysant lui même son découpage. Dès lors, l'ARN passa pour la première macromolécule biologique apparue sur Terre.
Il présente plusieurs intérêts: cette molécule possède de par sa structure une activité sur les protéines qui s'exerce actuellement dans des assemblages complexes, les ribosomes, dans lesquels l'ARN catalyse la formation de liaisons peptidiques entre acides aminés. D'autres ARN peuvent découper des protéines. De plus, ces molécules possèdent une certaine capacité d'évolution en réponse à leur environnement: l'ARN peut ainsi devenir résistant à la ribonucléase qui, normalement, le clive.
Bien que l'usage des polymères d'acides nucléiques soit aujourd'hui généralisé chez les êtres vivants, rien n'assure qu'il en a toujours été ainsi. Il est en effet difficile d'obtenir un ARN en milieu prébiotique, le seul acide nucléique facilement synthétisable étant l'adénine. De surcroît, la réplication de l'ARN nécessite des intermédiaires liés à des phosphates, ce qui est difficilement reproductible sans enzymes... Un autre problème est posé par la faible probabilité de la réplication moléculaire en milieu liquide, les nucléotides ne demandant qu'à se disperser au lieu de se disposer bien sagement sur les brins modèles de la molécule. Il semble bien que l'ARN soit encore une molécule trop évoluée, et qu'il a dû dériver d'un ancêtre plus simple. Malgré cela, cet acide nucléique doit avoir été impliqué assez rapidement dans les processus vitaux.
· L'information génétique a pu être de nature électromagnétique: ce sont les chercheurs C. Woese et G. Wachtershauser qui sont à l'origine de cette conception faisant intervenir, à des degrés divers, la reproduction des cristaux comme base de la reproduction du vivant. Nous verrons que cette approche est de loin la plus intéressante, car comme le déclare G Wachtershauser "on créée de l'ordre (biologique) à partir de l'ordre (cristallin) et non du chaos". La répartition des charges électriques dans les minéraux aurait orienté la synthèse des premières molécules, et comme cette répartition est conservée, avec certaines variations, lorsque le cristal se développe, cela fournit un support physique pour une reproduction primitive.
· Le support de l'information génétique n'a que peu d'importance, ce qui compte, c'est la forme. Je dirais que cette hypothèse, personnelle, pourrait se décliner sous le nom de "bande des trois": cristaux, protéines et acides nucléiques sont capables de transmettre des formes. Je fais donc l'hypothèse qu'ils ont travaillé ensemble pour reproduire les premières formes paravivantes: les cristaux fournissent un milieu capable de se propager, de se reproduire avec variation, entraînant un mélange d'acides nucléiques et aminés liés à sa surface. Nous verrons que les cristaux en question peuvent être ceux contenus dans les grains de poussière interstellaire, et que le début du processus vital, si ce n'est la totalité, a donc pu se dérouler dans l'espace.
Une hypothèse iconoclaste: le tout protéine
La majorité des chercheurs considèrent que l'ARN est la première molécule organique originelle. Le problème, c'est qu'en Science (comme ailleurs, mais ceci est une autre histoire) on ne doit (ou plutôt ne devrait) pas se prévaloir du nombre pour affirmer la véracité d'un propos, mais uniquement des faits expérimentaux: la vérité n'est pas démocratique. D'où cette hypothèse: et si nous avions eu au début de la vie une information protéique, bien plus simple, se transmettant simplement par contact avec des oligonucléotides? Cette idée aurait paru saugrenue il y a peu, jusqu'à la découverte du fameux prion, cette protéine extrêmement résistante (hélas!) capable de transmettre son "information" sans matériel nucléotidique.... La première molécule autoréplicative n'aurait elle pas pu être un "paléoprion", transmettant uniquement une forme dont découlerait, plus tard, une fonction... c'est une question ouverte.
A présent que nous avons vu quelles étaient les molécules des êtres vivants, voyons quelles sont les diverses théories expliquant leur origine. Nous verrons si elles sont capables de fournir un cadre cohérent permettant d'expliquer l'élaboration de la machinerie moléculaire qui équipe les êtres vivants actuels, même si cette dernière ne donne qu'une image déformée par des milliards d'années d'évolution de ce qu'elle était à l'origine.
Il existe deux grandes "chapelles" dans la biologie contemporaine, qui différent par leur approche du problème. Plus que les détails de leurs arguments, ce qui les oppose tient surtout à des conceptions quasi philosophiques: aux tenants d'une origine purement endogène de la vie, qu'ils limitent bien souvent à la Terre, s'opposent les zélateurs d'une origine essentiellement exogène, dont je suis, qui eux sont favorables à une large dispersion des formes vivantes.
Les frontières entre ces deux conceptions sont cependant mouvantes, et bien des conversions inopinées se produisent au gré des découvertes! Ainsi, après une époque où la vie n'était envisagée que sous l'angle terrestre, plusieurs indices ont renforcé le camp des tenant de l'origine extraterrestre. Nous verrons que certaines conceptions, loin de s'exclure mutuellement, peuvent utilement se compléter pour peu que l'on adopte un point de vue plus global. Ajoutons également l'existence de quelques "églises" dissidentes, tenues par des "intégristes" défendant des positions variées allant d'une vie uniquement terrestre d'obédience divine à l'influence d'éventuelles intelligences extra-terrestres sur son apparition et son développement, sans oublier l'interprétation au pied de la lettre de différents textes sacrés... Essayons de nous repérer et de poser des jalons dans ce foisonnement.
En 1950, le chimiste Harold Hurey propose une atmosphère primitive réductrice, riche en méthane et en ammoniac. Il se fonde pour cela sur l'étude de météorites et de l'atmosphère des planètes géantes. Trois ans plus tard, Stanley Miller a l'idée de reconstituer en laboratoire l'environnement de la Terre primitive. Sans se vanter de son idée, il bricole un montage expérimental en circuit fermé contenant de l'eau, les différents gaz de l'atmosphère et une source d'énergie, mimant les éclairs des orages de l'atmosphère primitive. Il fait circuler son mélange, préalablement stérilisé, afin de reproduire les phénomènes d'évaporation et de condensation. On se serait attendu à conserver uniquement les molécules que l'on y avait mises, mais il eu la surprise de découvrir que des dépôts bruns se formaient sur la paroi de son appareil. En analysant cette substance, il découvrit qu'il s'agissait d'acides aminés: ces briques à protéines s'étaient formées sous l'influence de la lumière et de la chaleur dans une reconstitution de l'océan primitif! Cette expérience eut un grand retentissement, car elle montrait que les molécules du vivant pouvait bien, expérimentalement, être récréées dans les conditions que l'on pensait être celles de la Terre primitive.
Beaucoup d'opposants à l'idée d'une formation "facile" de la vie à partir de molécules prébiotiques simples se sont arrêtés à cette expérience, et n'en ont pas suivi les développements ultérieurs. Certains de leurs arguments tombent ainsi d'eux-mêmes. L'expérience de Miller a été refaite bien des fois, en variant l'atmosphère de départ, l'intensité de la source d'énergie... mais l'on obtient toujours les acides aminés. A la fin des années 50, S. Fox montre que ces derniers s'organisent en petites sphères, les proténoïdes. Cependant, ces sphérules ne peuvent se reproduire, donc évoluer.
Actuellement, de nombreux biochimistes (Meli, Vergne, Maurel....) étudient des molécules synthétisées dans des conditions prébiotiques et qui font le lien entre la capacités informationelles de l'ARN et l'activité enzymatique des protéines. Ainsi, le N6 ribosyl-adénine est un analogue potentiel de l'acide aminé histidine, dont il reproduit le comportement chimique. D'autres dérivés N6 ou N3-ribosyl-purine ont pu être des maillons essentiels entre le monde des protéines et le monde de l'ARN. Comme nous le verrons par la suite, des ARNs se liant à des acides aminés ont pu servir d'ancêtres aux ARNt actuels, participant ainsi à l'émergence d'un code génétique.
|
Nous vivons dans un monde dont l'atmosphère contient 20 % de dioxygène, un gaz corrosif dont les êtres vivants se protègent par des dispositifs variés. Par exemple, des enzymes comme la catalase, la peroxydase ou la superoxyde dismutase permettent au métabolisme de résister à l'action délétère de l'oxygène et de ses dérivés. Pour les bactéries anaérobies qui ne disposent pas de ces protections, l'oxygène est toujours un poison mortel. Sans oxygène, pas d'ozone pour protéger la surface de la planète des U.V. solaires qui arrivent au sol, apportant l'énergie nécessaire aux synthèses chimiques à partir des éléments minéraux disponibles. Nombreux sont les indices qui montrent que l'oxygène était absent de l'atmosphère primitive, au début de la vie: - des dépôts d'uraninite UO2 datant de cette époque sont constitués. Or, ce minéral est dégradé en U3O8 soluble si la teneur en O2 est supérieure à 1%. - des couches rouges d'hématite Fe2O3 nous indiquent une teneur croissante en oxygène avec le temps. De tels dépôts d'hématite ont été identifiés sur Mars, renforçant encore l'hypothèse selon laquelle cette planète et la nôtre ont évolué de concert après leur formation. - des formations de fer rubané se sont déposées lorsque la teneur en O2 a augmenté, transformant le Fe2+ soluble en Fe3O4 insoluble qui a précipité avec de la silice. - le métabolisme des êtres vivants garde la trace de cette époque anoxique, toutes les réactions chimiques de base se déroulant en l'absence d'oxygène. |
Les réactions énergétiques qui permettent aux êtres vivants de récupérer de l'énergie à partir des molécules de leur alimentation sont de 4 types différents:
Les rééditions multiples de l'expérience de Miller conduites entre 1960 et 1980 nous ont montré également que des acides nucléiques peuvent se former dans la "soupe primitive" de Miller et s'accumuler. On se trouve cependant confronté à un problème: nous avons vu que les synthèses des protéines et des molécules d'ADN sont interdépendantes: comment les acides nucléiques peuvent-ils se reproduire sans les protéines? Et d'où viennent les protéines s'il n'y a pas d'acides nucléiques pour les produire? On se retrouvait alors dans la situation inconfortable de savoir qui, de la poule protéine ou de l'oeuf ADN, était né le premier...
Une autre difficulté allait se révéler: l'atmosphère primitive, nous l'avons vu, n'avait pas la composition prévue par Urey: elle était bien plus riche en gaz carbonique et en azote. Bien qu'une atmosphère réductrice aurait pu s'établir sous l'influence du rayonnement solaire, la température et la pression à la surface de la Terre devaient être bien plus élevées que ne le pensait Miller. Dans ses conditions, la synthèse de molécules prébiotiques autres que quelques acides aminés devient beaucoup plus complexe. Les acides nucléiques, en particulier, sont difficiles à obtenir.
Les chercheurs demeuraient donc dans l'expectative lorsque, en 1980, le biochimiste T. Cech démontre en étudiant un protozoaire que l'Acide Ribo Nucléique (ARN) est capable de se réparer et de se recopier seul, sans l'intervention de protéines: il est à la fois gène et enzyme. Du coup, il semble bien que l'on tienne un élément de réponse à notre dilemme: c'est "l'oeuf" ARN qui serait apparu en premier. Cette idée est présentée en 1986 par W. Gilbert sous le nom de "monde d'ARN": Les premiers organismes auraient été de simples molécules d'ARN autoreproducteurs. Secondairement, ils se seraient aidés de molécules de protéines pour se reproduire plus facilement. Cet ensemble ARN+protéines s'entoure ensuite d'une membrane lipidique qui concentre et protège ces molécules. Par la suite, l'ARN est remplacé par de l'ADN, plus stable.
Il ne fait plus de doute aujourd'hui qu'en effet l'ADN a succédé à l'ARN chez les êtres vivants: biochimiquement, l'ADN est un ARN modifié, et nos cellules effectuent toujours cette transformation. Le principal problème, c'est justement d'obtenir cet ARN de départ! La formation spontanée, en solution (nous verrons l'importance de ce terme) de cette molécule est assez peu probable, mais reste possible. C'est cependant faire une hypothèse risquée de croire que les molécules autoreproductrices primitives ressemblaient aux nôtres. Une autre molécule, plus simple que l'ARN, pourrait avoir joué son rôle, c'est du moins ce que pense L. Orgel, de l'institut Salk. En effet, la capacité à s'autoreproduire ne nécessite pas obligatoirement une grande complexité: le chimiste J. Rebek a obtenu expérimentalement des molécules de synthèse capables de s'autoreproduire, mais qui sont bien plus simples que le moindre brin d'ARN! De plus, l'ARN est une molécule fragile. Initialement, on pensait que cette molécule, apparue en solution, s'était concentrée petit à petit dans des petites mares tièdes... nous avons vu que les conditions de la Terre primitive ne permettent pas de tels processus.
Bien que l'ARN soit très probablement impliqué dans l'origine de la vie, il ne peut, seul, être à la base de celle-ci. Actuellement J Szostak et M. Eigen ont montré que l'ARN permettait toutes les réactions chimiques caractérisant un organisme, et l'on cherche comment cette molécule a pu s'entourer d'une membrane. Mais l'hypothèse d'ARN ancestraux de même structure chimique que les nôtres apparaît de plus en plus risquée: les molécules évoluent, et plus le temps dont elle dispose est grand et plus cette évolution est intense, surtout si la pression de sélection, augmentée encore par l'environnement primitif, accélère encore cette évolution. Ainsi, ces dernières années, P. Forterre et S. Doolittle ont signalé que, aux vues des spécificités biochimiques et génomiques des eucaryotes, des bactéries et des archaebactéries, la théorie d'un ancêtre unique des formes vivantes est sans doute erronée: la vie serait apparue plusieurs fois, dans des environnements et des conditions différentes, et ces formes de vie primaires différentes évoluant simultanément se seraient enrichies mutuellement par des transferts de gènes. Certaines auraient disparu, trois au moins seraient à l'origine du monde vivant terrestre.
Si l'on conserve l'hypothèse d'un ARN primitif, une controverse existe quant aux conditions thermiques du milieu où cette molécule a été synthétisée. Comme on pensait que les formes de vie les plus primitives étaient les archaebactéries vivant dans des milieux chauds, on a conjecturé que l'ARN avait une origine impliquant des hautes températures. En 1999, l'étude des ARN ribosomaux montre que ces derniers ne contiennent pas assez de guanine (G) et cytosine (C) pour avoir résisté aux hautes températures (54% au lieu de 60% minimum). En effet, G et C sont liés par 3 liaisons alors que les deux autres bases ne le sont que par deux. Il en résulte qu'un milieu chaud va sélectionner les liaisons les plus solides (3) qui devront être plus nombreuses que les liaisons à 2. Le dernier ancêtre commun (si tant est qu'il y en ait eu un et un seul!) devait donc vivre en milieu tempéré. Cependant, un réexamen des conditions de cette étude ainsi que d'autres considérations laissent entendre que l'ARN ancestral prend bien son origine dans un milieu chaud. Voilà qui est intéressant vu que toute la surface terrestre était "un milieu chaud!" (on suppose une température de 80°C au sol!). Cette controverse laisse entendre que l'idée d'une synthèse d'ARN autoreproducteurs en solution constituant la base à partir de laquelle l'évolution moléculaire conduit aux êtres vivants est sans doute erronée, car cette molécule n'aurait pas pu se former de cette façon dans les conditions de la Terre primitive, il y a... 4 milliards d'années.
Avant d'examiner d'autres idées, revenons un moment sur l'expérience historique de Miller. Nous avons vu que le mélange gazeux qu'il a utilisé n'était pas adéquat pour décrire l'atmosphère terrestre ancestrale. Cependant, les résultats que lui et ses successeurs ont obtenus sont indéniables. Étrangement, personne à ma connaissance n'a fait remarquer que si l'atmosphère de Miller ne s'appliquait pas à la Terre, elle convenait par contre parfaitement à la description des atmosphères des planètes joviennes comme Jupiter et Saturne: les enveloppes gazeuses de ces dernières contiennent méthane, ammoniac et vapeur d'eau. L'énergie n'y manque pas, par exemple à cause d'aurores boréales intenses, et d'orages gigantesques. Lorsque l'on s'enfonce dans leurs couches nuageuses, la température et la pression augmentent graduellement. On en déduit que l'on se retrouve dans les conditions de l'expérience de Miller, et que fatalement des acides aminés et nucléiques ont dû se former dans cette énorme atmosphère. Dès lors, des formes de vie bactériennes, vivant en suspension dans l'atmosphère auraient très bien pu se développer. Sur notre propre planète, de telles bactéries atmosphériques ont récemment été mises en évidence. Sur Jupiter, les forces de gravité sont telles (3 fois la gravité terrestre) qu'elles ont pu interdire l'apparition de formes de vie trop massives, celles-ci étant entraînées vers le centre de la planète, à des pressions et des températures incompatibles avec le maintien de structures moléculaires complexes. Une vie bactérienne dans les atmosphères des planètes joviennes est en accord avec tout ce que l'on sait de l'apparition de la vie, mais elle n'a jamais été considérée avec sérieux, se heurtant à un obstacle épistémologique: comme nous habitons une planète tellurique, nous voulons à toute force que la vie soit limitée à ce type planétaire. Il n'en est probablement rien, et qui sait si les délicates couleurs des nuages de Jupiter ne proviennent pas de molécules rejetées par des bactéries joviennes? Si cela était le cas, nous pourrions bien observer depuis des siècles, même dans les plus petits télescopes, la manifestation de formes de vie que nous cherchons partout ailleurs alors qu'elle nous crève les yeux... Cela est selon moi d'autant plus probable que, comme nous le verrons, même si les planètes joviennes n'abritent pas de vie endogène elles ont du cependant recevoir des "voyageurs de l'espace" microbiens en provenance de... la Terre ou Mars.
Les difficultés à trouver un environnement suffisamment protégé pour que les premières formes de vie résistent aux influences destructrices des rayonnements et des chutes de météores ont conduit à considérer un environnement que, de prime abord, on avait cru totalement impropre à la vie: les abysses. Ces fonds marins ne recevant aucune lumière solaire et constituant un environnement où la pression hydrostatique est énorme, on les avait cru sans vie jusqu'à ce que les premiers bathyscaphes révèlent l'existence d'animaux à ces grandes profondeurs. En 1977, J Corliss a découvert des formes de vie dans ce que l'on prenait pour un désert: à 2500 m de fond sous l'océan, près de sources hydrothermales chaudes, vivent des vers tubicoles géants (2m), les riftia. Ils supportent une pression de 250 atmosphères, des températures de plusieurs centaines de degrés, un environnement acide et riche en gaz "toxique". (Un sujet de réflexion pour tous ceux qui déclarent péremptoirement un milieu "impropre à la vie" parce qu'il n'est pas assez "confortable"). Ces vers vivent en symbiose avec des bactéries qui tirent leur énergie des composés soufrés "toxiques" émis par les sources prenant leur origine dans le magma terrestre. D'autres animaux (des mollusques comme Calyptogena et Bathymodiolus) vivent dans les mêmes niches écologiques que les riftia et utilisent les mêmes bactéries. Corliss propose que les premières formes de vie se soient formées en profondeur, près de ces sources chaudes. Elles auraient tiré leur énergie des gaz produits à ce niveau. Leur grande profondeur les aurait protégé du rayonnement solaire et des impacts météoritiques. Cette hypothèse semble corroborée par C. Woese, qui en étudiant le matériel génétique d'unicellulaires a mis en évidence l'existence des archaebactéries. Ces dernières tirent leur énergie de composés chimiques dissous, résistent à de fortes températures et à un environnement semblable à celui des sources "fumeurs noirs" où vivent les riftia. Progressivement, des analyses génétiques ont situé l'ancêtre commun des cellules chez les archaebactéries. La vie serait donc apparue au niveau des "fumeurs noirs", dans un milieu très chaud et sous forte pression.
Cette conception cadre parfaitement avec ce que l'on sait de l'arbre généalogique des cellules, et nous avons vu que les conditions physiques à l'époque de l'apparition de la vie militent en faveur d'organismes primitifs apparus à des températures proches de 100°C. Il reste cependant quelques zones d'ombre dont la plus étendue concerne les acides nucléiques: il est a peu près sûr que l'ARN est apparu avant l'ADN. Mais l'ARN est détruit facilement à haute température, alors que l'ADN y résiste! C'est pour cela que le chercheur P. Forterre remet en question cette hypothèse. Cependant, une origine "chaude" de la vie reste probable puisque les régions "froides" semblent avoir été plus que rares à l'époque... Le problème des acides nucléiques allait cependant être repris et développé par un chercheur original dont nous reparlerons plus en détail, G. Wachtershauser.
Une autre difficulté est la durée d'activité des sources hydrothermales: elle n'est actuellement que de quelques années avant que ces sources de nutriments se tarissent: rien n'indique que cette durée soit suffisante pour permettre l'apparition des premières formes de vie, sauf à admettre que leur développement ait été bien plus rapide qu'on ne le croit généralement.
Nos ancêtres les minéraux : une "aurore de pierre" à l'aube de la vie
La formation des premiers acides nucléiques posant problème, un chimiste travaillant au bureau des brevets, à Berne, et occupant la même fonction qu'un certain Albert Einstein, reconsidère le problème de l'émergence du vivant en proposant que la première manifestation de la vie n'est pas l'autoreproduction mais l'établissement d'un métabolisme capable de capter du carbone et d'utiliser de l'énergie: en 1988, Gunter Wachtershauser imagine ainsi une origine minérale de la vie. A la surface de cristaux de pyrite, des molécules organiques se fixent pour former des "métabolistes" qui tirent leur énergie du sulfure de fer. Ils produisent ainsi de la matière organique à partir de minéraux. La croissance de cristaux de pyrite fournit une division primitive qui permet la reproduction du "métaboliste" par divisions successives. Ce métabolisme primitif utilise des composés soufrés, dont C. de Duve montre l'intérêt et l'utilisation par certains organismes pour produire les acides ribonucléiques, et donc l'ARN. Cette voie de synthèse est la seule actuellement plausible.
Antérieurement, A. Cairns-Smith avait proposé une théorie similaire mais basée sur l'argile dont les cristaux permettent une réplication avec erreurs, donc la possibilité d'une évolution. Si ce chercheur envisageait un support minéral soumis à des cycles d'asséchement et d'hydratation, et bordant donc une mer primitive, ou des cours d'eau; G. Wachtershauser montre que les synthèses de ses "métabolistes" réclament chaleur, forte pression et disponibilité abondante de minéraux soufrés: ces conditions correspondent à celles régnant près des sources hydrothermales profondes. Il situe donc l'origine de la vie au niveau des sources hydrothermales des abysses, tout comme Corliss. Cependant, d'autres milieux peuvent avoir présenté des caractères comparables: intérieur des roches terrestres, surface sous forte pression atmosphérique avec éruptions volcaniques ou geysers...
L'idée que les minéraux ait leur mot à dire sur l'origine des premières formes de vie est extrêmement importante: elle permet, nous le verrons, d'expliquer pourquoi les premières formes de vie ne se sont pas dissoutes dans la "soupe primitive" dont l'existence devient de plus en plus suspecte. Nous verrons que le lien entre les mondes minéral et organique prend forme, selon moi, dans les nuages de poussière interstellaire, et ne fait par la suite que se renforcer jusqu'à l'émergence du vivant. La récente mise en évidence par Folk puis Uwins de l'existence de nano-organismes liés aux processus de minéralisation vient confirmer de façon éclatante le bien fondé de cette conception.
Ce sont les faits et les arguments de cette communauté de pensée qui m'ont conduit à publier ces pages. S'ils conduisent parfois à des excès préjudiciables à sa valeur scientifique, ils ont l'avantage de nous fournir les bases d'une nouvelle révolution copernicienne visant à établir la banalité du vivant.
Ce sont les kangourous qui ont été surpris! Le dimanche 28 Septembre 1969, un éclair lumineux zébra le ciel de l'Australie, près de la ville de Murchison. On récupéra au sol une météorite de 225 kg, dont l'analyse allait révéler bien des surprises: elle montra que cette roche contenait dans des inclusions noirâtres appelées chondres 17 acides aminés différents. Cette météorite ne provient pas de la fragmentation d'un corps plus grand mais s'est formée en même temps que le système solaire, et les rapports isotopiques de certains éléments, comme le néon, nous montrent en outre qu'elle provient d'une région voisine d'étoiles explosives du type Nova ou Supernovae. Nous devons en conclure contre toute attente que les acides aminés peuvent se former dans le milieu interstellaire. Des météores plus anciens, étudiés avec les techniques modernes, ont eux aussi révélé contenir des acides aminés. 8 acides aminés sur les 20 utilisés par les êtres vivants ont été retrouvés dans des météorites. De plus, certains de ces acides aminés montrent l'existence d'une dissymétrie entre leurs formes chimiques "droite" D et "gauche" L, ce qui est constaté chez les êtres vivants, qui n'utilisent que des formes L.

De nombreuses molécules organiques (plus de 500 !) ont été identifiées dans la météorite de Murchison: en décembre 2001, l'équipe de G. Cooper a annoncé la détection d'un sucre simple et de polyols comme le glycérol. Auparavant, à partir des composés extraits de cette roche, D. Deamer avait obtenu des membranes constituées de molécules organiques. Ces vésicules délimitent un milieu qui serait favorable à la concentration de molécules prébiotiques.
Des expériences réalisées en orbite terrestre (programme "biopan") ont montré qu'il suffisait d'une couche de 5 microns de poussière météoritique pour protéger les acides aminés de la destruction. Même des météorites de toute petite taille (+ de 5 microns...) sont donc capables d'avoir transporté ces molécules. Ce point est important, car les micrométéorites sont à la fois extrémement abondantes et particulièrement résistantes: plus de 80 % d'entre elles ne fondent pas lorsqu'elles traversent notre atmosphère, la combustion d'une partie du carbone contenue dans ces météores provoquant une détente gazeuse refroidissant le grain et lui assurant ainsi une "climatisation" protectrice pendant sa chute. Chaque année, ces météores apportent 20 t de carbone extraterrestre sur notre planète, ce qui représente un apport hallucinant de 1017 tonnes de C depuis la formation de notre planète, soit 5 ordres de grandeur au dessus de la quantité de carbone contenue dans l'ensemble des êtres vivants!
Parmi les acides aminés d'origine extraterrestre, certains sont inconnus sur Terre. Nous avons vu que les êtres vivants n'utilisent qu'une vingtaine d'AA différents, alors qu'il en existe plusieurs centaines de possibles. A la publication des résultats obtenus sur ces acides aminés, certains invoquèrent une contamination de l'intérieur du météore par des molécules terrestres, mais la présence de ces acides aminés inconnus donna un argument fort en faveur de l'existence de processus de synthèse d'acides aminés en milieu interstellaire. Cependant, ces molécules "étrangères" furent pendant presque 20 ans considérées comme quantité négligeable dans l'histoire de la vie, comme des curiosités de laboratoire sans rapport avec l'histoire de la vie ici bas. Seul les astrophysiciens Hoyle et Wickramasinghe insistèrent sur l'importance de ces apports exogènes, mais leurs exagérations ainsi que le caractère révolutionnaire de leurs conceptions les desservirent, et leur apport, venu trop tôt, resta lettre morte.
En effet, après la vague "écologiste" des années 60 et les photos splendides de la Terre flottant dans l'espace, un courant d'idée "isolationniste" traversa les sciences en général et la biologie en particulier. L'idée que nous "n'avions que la Terre" et qu'il fallait la protéger déboucha, inconsciemment, sur la nécessité de décrire toute l'histoire terrestre sans faire appel à des influences extérieures, mal connues des géologues et biologistes et considérées comme des relents d'anciennes conceptions catastrophistes où se mêlaient les mythes religieux et les premiers travaux des géologues, au début du 19éme siècle. Ainsi, il ne faut jamais oublier que la science est faite par des hommes et des femmes sensibles, parfois à leur insu, à l'air du temps, aux modes qui peuvent faire accepter ou rejeter une idée indépendamment de sa valeur scientifique. Le cloisonnement de la recherche ainsi que l'absence d'une véritable culture scientifique chez la plupart des chercheurs les poussent à dénigrer systématiquement ce qui ne constitue pas l'objet de leurs recherches: biologistes et géologues, unis pour découvrir l'origine de la vie, considéraient alors fermement qu'ils ne devaient s'occuper que de ce qui s'était passé sur Terre seulement, le reste étant considéré comme étant sans intérêt.
Cette attitude fut modifiée pendant les années 80, lorsqu'il fallu expliquer les extinctions en masse qui jalonnent l'histoire de la vie, ces périodes où, en quelques milliers d'années, entre 75 et 90% des espèces vivantes disparaissent de la surface de la Terre pour être ensuite remplacées par de nouvelles espèces... Le géologue Walter Alvarez, de l'université de Berkeley, découvrit en 1980 une fine couche d'argile marquant la limite entre le crétacé et le paléocéne. L'analyse de cette couche, retrouvée partout dans le monde, et qui affleure en France sur la côte Basque, montra qu'elle était singulièrement enrichie en un métal extrêmement rare sur Terre mais plus commun dans les météorites, l'iridium. Par un hasard bienheureux, le père du géologue Alvarez n'était autre que le physicien (et prix Nobel) Luis Alvarez, spécialiste des réactions nucléaires de plein air (bombes etc...) qui avait étudié les effets d'explosions violentes pour le département de la défense US. Il avait mis en évidence qu'une guerre atomique, ou un impact de météorite, aurait pour effet de mettre en suspension dans l'atmosphère assez de poussières pour provoquer un "hiver nucléaire" qui interromprait la photosynthèse des plantes et détruirait nombre de formes de vie sur Terre. Dès lors, Pour Alvarez junior, la cause était entendue: un vilain météore avait percuté la Terre à la limite crétacé/tertiaire et avait précipité la fin des célébrissimes dinosaures. Outre l'iridium excédentaire, d'autres indices d'un impact majeur furent découverts: des sphérules basaltiques, fragments de croûte océanique fondue sous l'impact, des grains de quartz ayant subi des déformations caractéristiques des cratères météoritiques et des minéraux comme le sélénium, le rhénium, l'arsenic, le sélénium et l'antimoine. Les conclusions d'Alvarez furent loin d'être acceptées, et sont même encore combattues par des géologues qui veulent à toute force que seules des origines endogènes, comme des éruptions volcaniques massives, suffisent à expliquer cette extinction. Leur principal argument était que le cratère d'impact n'avait pas été trouvé. Il avait été pourtant mis en évidence par des mesures sismiques dès 1970, mais il fallu un an de recherche pour le retrouver dans la péninsule du Yucatán, à l'est du Mexique, à 200 km au large du village de Puerto Chicxulub: ce cratère est aujourd'hui rempli de sédiments et possède un diamètre de 200 km. Sa profondeur atteint 20 km. D'autres travaux ont montré par ailleurs qu'un impact météoritique majeur était capable de générer, à très longue distance, des éruptions volcaniques intenses, apportant du baume au coeur des tenants des extinctions par éruptions volcaniques massives.
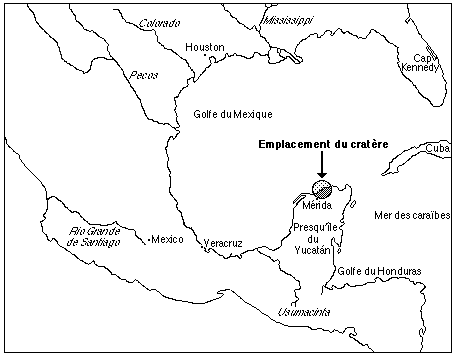
Cette origine météoritique de la fin de ces animaux emblématiques que furent les dinosaures amena un regain d'intérêt pour l'environnement cosmique de notre planète. Le cinéma s'en fit l'écho, et quelques films et téléfilms de l'époque achevèrent de populariser les effets des impacts météoritiques. Cette prise de conscience de l'importance de notre environnement spatial déboucha sur plusieurs découvertes mettant en évidence son rôle fondamental non seulement dans l'apparition mais aussi dans le développement de la vie sur notre planète.
| Ci contre, une carotte exposée au Smithsonian museum of natural history, à Washington DC, montrant la transition entre l'époque des dinosaures ((flèche jaune) et celle des mammifères (en rouge). Entre les deux, la couche sombre contient des matériaux d'origine extraterrestre. |  |
Ainsi, en 1989, une couche d'argile datant de la fin du crétacé se révéla contenir des acides aminés inconnus chez les animaux. K. Zahnle et D. Grinspoon suggérèrent alors, 10 ans après Hoyle, que ces molécules organiques pouvaient être d'origine cométaire. Les comètes et/ou les météorites ont donc pu apporter des éléments organiques sur la Terre primitive. Mais, direz-vous, ces corps s'échauffent fortement en traversant l'atmosphère, et les molécules organiques qu'ils contiennent doivent être détruites. Ce n'est pas le cas pour les corps possédant une masse relativement faible, de l'ordre de quelques dizaines de kg: la vitesse finale de chute n'est alors "que" de 200 m/s environ car l'air a totalement freiné le météore qui tombe alors en chute libre. L'échauffement de la roche se limite alors à une couche externe d'une épaisseur variant de quelques mm à quelques cm, le coeur restant froid (la majorité de l'énergie produite servant à chauffer l'air). De plus, l'ablation des météorites a pu provoquer des réactions chimiques prébiotiques dans l'atmosphère, et disperser dans l'air des poussières contenant des composés organiques, comme l'a montré C. Chyba. Il faut cependant voir que l'essentiel des matériaux extraterrestres qui parviennent sur notre planète ne le font plus sous la forme de spectaculaires et dangereuses collisions: outre les météores et les noyaux cométaires, l'essentiel des contributions exogènes à l'apparition de la vie pourrait bien avoir été constitué par des particules de poussière interstellaire, aussi petites, sinon plus, que celles de la fumée de cigarette (Dieu serait-il bien un fumeur de Havanes?), qui pénètrent dans notre atmosphère sans être échauffées du fait de leur faible masse et de leur vitesse limitée. Nous examinerons plus en détail dans le chapitre suivant dans quelle mesure ces grains de poussière ont pu constituer le support minéral sur lequel les premiers organismes vivants ont pu se développer.
L'étude des météorites allait livrer des informations encore plus sensationnelles. En effet, en 1998, le Pr. Folk identifia dans la météorite Murchison des formations pouvant être les reliquats de nano-organismes fossilisés. Des formations similaires ont également été identifiées dans d'autres météorites (Allende, ALH 84001, Tataouine). Bien entendu, la controverse fait rage autour de l'interprétation des structures observées: certains y voient les fossiles de formes de vie extraterrestres, d'autres de simples artefacts (des "mirages"), d'autres encore des contaminations purement terrestres. Le débat est loin d'être clos, ce qu'illustre fort bien la controverse concernant la météorite ALH 84001.
Ce caillou récupéré dans l'antarctique s'est révélé, de par la composition isotopique des gaz piégés dans sa masse, provenir de la planète Mars, tout comme 12 autres météores que l'on désigne par l'acronyme SNC (faisant référence aux sites de découverte des 3 premières d'entre elles, Shergotty, Nakhla et Chassigny). En 1996, Mc Kay analyse ALH 84001 et pense mettre en évidence l'existence de matière organique martienne. Il interprète les microstructures observées dans cette météorite comme des microfossiles bactériens, première preuve directe de l'existence d'une vie sur Mars. Cependant, l'interprétation de ces structures est ardue, et d'autres hypothèses non biologiques peuvent les expliquer. La petite taille des "fossiles" observés, présentée un temps comme s'opposant à leur origine biologique, n'est plus un argument valable depuis la découverte (toujours âprement discutée) des nanobactéries. Notons qu'une autre météorite martienne, EETA 79001, a libéré après chauffage du dioxyde de carbone enrichi en 12C. Cet enrichissement isotopique est, lorsqu'il est détecté sur Terre, interprété comme étant d'origine biologique. Une contamination terrestre étant toujours possible, il faudra, pour en savoir plus, prendre la direction des formations sédimentaires identifiées récemment à la surface de la planète rouge.
Abordons maintenant une hypothèse qui découle un peu de la précédente, et dont l'aspect "science fiction" ne doit pas masquer l'intérêt. Son idée maîtresse est que la vie n'est pas apparue sur Terre, mais y a été apportée. Cet apport, pour les défenseurs les plus raisonnables de cette thèse, a été involontaire, alors que d'autres, et non des moindres, font intervenir un ensemencement volontaire de la planète. Remarquons tout d'abord que le problème n'est alors que déplacé, car il faut expliquer l'origine de cette vie exogène. Ensuite, je dois avouer que j'ai eu moi-même pendant des années assez peu de considération pour cette hypothèse. Elle me semblait par trop fantastique, et avait été par le passé trop discréditée par nombre d'imposteurs et de délirants archéomânes pour conserver de l'intérêt. C'est l'étude des conditions générales de l'apparition de la vie et des découvertes les plus récentes qui m'a conduit à modifier ma conception et à tenir comme très probable le côté le plus raisonnable de cette thèse. Le seul problème étant de mettre en balance ce qui, chez les premiers êtres vivants, était spécifiquement endogène, terrestre, et ce qui pouvait provenir du milieu interplanétaire, voire interstellaire.
Dès 1890, le chimiste S. Arrhenius avait proposé que des micro-organismes dérivant dans l'espace aient colonisé la Terre. Cette idée resta cependant isolée, et les progrès de l'astronomie, ainsi que les difficultés de l'astronautique naissante, établirent que l'espace constituait bien un des milieux les plus hostiles à la vie. Dès lors, tous ceux qui osaient exprimer l'idée d'une vie dans l'espace étaient considérés au mieux comme de doux dingues, au pire comme des hérétiques auquel il fallait absolument barrer tout accès vers des postes universitaires... Encore aurait-il fallu s'entendre sur ce que l'on considérait comme formes de vie! Les premiers doutes se firent jour lorsque la récupération sur la Lune d'une caméra de la sonde automatique surveyor par les astronautes d'Apollo 12 montra qu'une bactérie au moins avait pu sans dommage faire le voyage Terre-Lune et demeurer dix ans sur cet astre. Dans les années 70, les microbiologistes allèrent de surprise en surprise, découvrant que les bactéries pouvaient pratiquement coloniser n'importe quel milieu: la découverte de ces formes de vie "extrémophiles" capables de résister à la chaleur, au froid, aux radiations, à l'absence d'eau... démontra que la vie microbienne était bien plus résistante que l'on ne l'avait cru.
Devant les difficultés à expliciter l'origine de la vie sur notre planète, l'hypothèse "alien" fit un retour en force. Certains utilisèrent leur autorité scientifique pour oser les hypothèses les plus folles, et c'est ainsi qu'en 1980 le prix Nobel F. Crick, co-découvreur avec T. Watson et R. Franklin de la structure de l'ADN, et L. Orgel avancèrent que la Terre et d'autres planètes avait été ensemencées volontairement par des intelligences extra-terrestres. Nous verrons que, sans aller aussi loin, l'hypothèse d'un ensemencement de la Terre par des micro-organismes ne peut être rejetée, ni même celle selon laquelle notre propre planète ait pu servir de "source de vie" non seulement pour le système solaire, mais peut être aussi pour une bonne part de la galaxie! Il est en effet de plus en plus probable que des bactéries puissent être arrachées à leur planète d'origine et demeurer vivantes des millions d'années avant d'atteindre d'autres corps célestes.
En 1990 L'inénarrable et regretté F. Hoyle, poil à gratter de l'astrophysique, soutint l'idée, qu'il développait depuis des années, selon laquelle non seulement des micro-organismes extraterrestres ont colonisé la Terre et y ont apporté la vie mais aussi selon laquelle cet apport exogène serait toujours actif, périodique, conférant ainsi une origine extraterrestre aux virus de la grippe ou du sida, et plus récemment du prion, agent causal de l'encéphalopathie spongiforme. Sans soutenir ces exagérations, fruits d'un isolement académique soigneusement entretenu autour des Pr Hoyle et Wickramasinghe (en toute justice, F. Hoyle par exemple aurait dû recevoir le prix Nobel pour ses travaux en astrophysique), force est de reconnaître que ces deux chercheurs ont dans le passé émis quelques idées très en avance sur leur temps, sur les molécules du milieu interstellaire par exemple, dont certaines qui furent confirmées par la suite. Nous verrons comment "faire le tri" entre ce qui est scientifiquement raisonnable et ce qui, pour être possible, reste assez improbable à ce jour.
Prendre en compte des apports extraterrestres de micro-organismes permettrait d'expliquer par exemple pourquoi plusieurs enzymes importantes du métabolisme ont besoin pour fonctionner d'un élément plutôt rare sur notre planète, le molybdène. Nous verrons cependant que ce problème est accessoire en comparaison d'autres que l'origine extraterrestre de la vie permet de résoudre.
A présent que nous avons passé en revue comment était organisé le milieu interstellaire, assisté à la formation du système solaire primitif, vu quels étaient les constituants et l'organisation générale des êtres vivants et quelles étaient les hypothèses se rapportant à leur apparition, nous sommes suffisamment armés pour essayer de proposer une vision synthétique qui, en utilisant les apports récents de l'astrophysique et de la microbiologie, va nous conduire à proposer une apparition générale et synchrone de la vie dans d'immenses régions de notre galaxie. Le fait qu'on ne puisse plus considérer la Terre comme étant une planète biologiquement isolée nous amènera à postuler l'existence d'une région isobiologique galactique, d'un réseau vital de taille astronomique au sein duquel les formes de vie, tout au moins microbiennes, sont similaires.