L'apoptose, un suicide
cellulaire aux effets salutaires
L'apoptose est une autodestruction cellulaire d'origine endogène. Ce mécanisme prend une grande importance dans l'explication des processus du développement, mais est également impliquées dans de nombreuses pathologies (cancers, SIDA) ainsi que dans des processus évolutifs.
Les cellules subissant une attaque exogène sont nécrosées: elles éclatent, provoquent la destruction de leurs voisines et déclenchent un processus inflammatoire suivi d'une cicatrisation impliquant une fibrose. Dans le cas de l'apoptose, la cellule se suicide: il n'y a ni inflammation, ni lésion des tissus. La cellule apoptotique se sépare de ses voisines puis se désintègre de façon ordonnée (condensation puis fragmentation du noyau, cassure régulière des chromosomes puis bourgeonnement du cytoplasme sous forme de petits ballonnets rapidement ingérés par les cellules voisines).
Il est apparu que dans l'organisme toute cellule est en état de mort imminente: elle ne survit que si elle reçoit de son environnement des signaux moléculaires qui inhibent son suicide. Ces signaux peuvent être de nature variés: outre des facteurs de croissance, des hormones sexuelles ou d'autres molécules peuvent inhiber ou déclencher le suicide cellulaire:
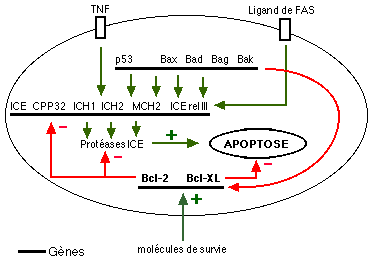
Originellement mis en évidence chez un vers nématode (Caenorhabditis elegans - R. Horvitz, 1986), les gènes qui commandent l'apoptose ont été retrouvés chez l'homme. Leur persistance et leur similarité chez des organismes aussi phylogénétiquement éloignés démontre leur importance. Leur action est schématisée sur la figure suivante:
Les gènes impliqués dans l'apoptose sont également mis en jeu dans de nombreuses pathologies: chaque fois qu'une population cellulaire meurt sans phénomène de nécrose ou prolifère exagérément, une implication de l'apoptose peut être envisagée.
|
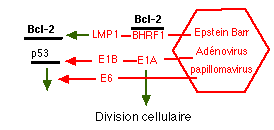
C'est le cas dans les cancers mais aussi dans les maladies neuro-dégénératives, dans certaines maladies virales et, nous l'allons voir, dans le diabète. Ci contre: Les virus cherchent à éviter le suicide des cellules qu'ils infectent en agissant directement sur les gènes de l'apoptose (les gènes viraux et leurs produits sont en rouge) |
 |
L'apoptose est caractérisée par des modifications du métabolisme cellulaires causant la destruction de la cellule. Elle peut donc être détectée par des modifications de la morphologie cellulaire ou de l'activité biochimique de la cellule:
|
|
|
|
|
Une autre forme de suicide cellulaire a été mis en évidence par Dale Bredesen (Buck Institute for Age Research) : la paraptose, au cours de laquelle des vacuoles se déceloppent à l'intirieur du cytoplasme, donnant à la cellule un aspect voisin mais différent de celui caractérisant une nécrose. Contrairement à l'apotose, la paraptose déclenche une réaction inflammatoire modérée. Les phénomènes papraptotiques étant présents chez des organismes très ancien comme des champignons, il est possible qu'elle représente une des premières voies de mort cellulaire programmée s'étant développée au cours de l'évolution.
Apoptose des cellules acineuses pancréatiques
Le suicide des cellules acineuses pancréatiques a été observé dans différents cas de figures mimant une pancréatite.Il peut être consécutif à:
Après la ligature du canal pancréatique, un oedéme instersticiel augmente le volume du pancréas qui décroît ensuite très vite. Le tissu exocrine s'atrophie à 90% en 7 jours chez la souris (Watanabe & al., 95) . Les cellules acineuses disparaissent par apoptose (Abe & al., 95), et de nombreux macrophages les ingèrent. En même temps, les cellules ductales prolifèrent pendant 2 semaines, alors que le pancréas reprend un volume normal en 8 semaine grâce à un développement de cellules adipeuses.
L'apoptose des cellules acineuses après une stress pancréatique est un phénomène général, qui conduit à de profonds remaniements tissulaires et cellulaires (Walker & al., 1992) dont le plus remarquable est le développement des cellules ductales et des cellules ovales. Néanmoins, l'involution du pancréas constatée après ligature du canal pancréatique fait également intervenir, dans des proportions et des prédominances variables selon les espèces, des phénomènes classiques de nécrose (Gukovskaya & al. - 1996). Les cellules acineuses sont cependant particulièrement "sensibles" à l'apoptose car le gène Bax s'exprime davantage que le gène "anti-apoptotique" Bcl2 dans le pancréas exocrine (Krajewsky & al., 1994). L'apoptose peut également se révéler bénéfique en limitant l'extension des dommages subis par les cellules acineuses au cour d'une pancréatite par exemple (Kaiser & al., 1996).
L 'apoptose des cellules acineuse observée par exemple après une nutrition sans cuivre est due à des perturbation de l'environnement et des interactions cellulaires. La disparition des cellules acineuses est compensée par le développement des cellules épithéliales des ductules et des cellules ovales. Ces cellules se trouvant dans un environnement différent deviennent des hépatocytes pancréatiques ayant les mêmes propriétés que ceux du foie (Rao & al., 1995)
Les effets de la ligature du canal
pancréatique sur les cellules ductales ainsi que leur
évolution ont été mis en évidence chez le
rat (Walker & al.,
1990, Rao & al., 1995) et particulièrement étudiés
chez le lapin par C. Cortie :
"Evolution des ductules et de leurs processus
prolifératifs chez le lapin
Extrait de la thèse du Dr. C. Cortie - "Evolution a long terme du syndrome diabétique non insulino dépendant chez le lapin: biochimie et histopathophysiologie - Institut National Polytechnique -Toulouse 14/04/1993
La ligature du canal pancréatique chez le lapin permet, comme chez le rat atteint de pancréatite chronique, de corréler à la fois le dépôt de collagène et le processus de destruction des cellules acineuses (Adler et al., 1979). Ces deux phénomènes sont concomitants avec des proliférations cellulaires.
Ainsi, dans les premiers stades post ligature chez le lapin , lorsque les cellules des acini ont involué, Petkov et al. (1992) ont rapporté la présence de corps denses, ce qui semblerait traduire une dégénérescence acineuse particulière de type apoptotique. En effet,Walker et al. (1992) précisent que l'élimination de ces cellules se fait non pas par nécrose mais le plus souvent par apoptose.La plupart des corps apoptotiques sont dégradés par des phagocytes mononucléaires qui résident entre les cellules épithéliales. Le mode de destruction de ces cellules acineuses peut être corrélé à la prolifération de cellules spécifiques.
Ceci se fait normalement sans rupture de la membrane basale et est accompagné simultanément par la prolifération des cellules centroacineuses. Ce même processus de prolifération a été rapporté chez le lapin, dès le 5ème jour post ligature (Petkov et al., 1992), et pourrait expliquer la variation d'épaisseur de l'épithélium des ductules observée nettement dès le 15ème jour sous forme de pseudostratification. Des travaux antérieurs (Catala et al., 1990) ont signalé chez ces animaux, au 30ème jour, une intensification des processus prolifératifs au niveau de l'épithélium ductal par une activité enzymatique élevée en particulier de la phosphatase acide (critère d'activité cellulaire).
Cependant, à partir du 180ème jour et de façon de plus en plus marquée au 450ème jour, pour un certain nombre de ductules, l'épithélium régresse et devient monocouche laissant supposer au contraire que les proliférations ont cessé, ce qui est nettement démontré par la dégénérescence de la plupart des ductules dont il ne reste que la membrane collapsée. Ceci est en accord avec les résultats décrits chez le rat après ligature où les phénomènes extensifs de prolifération de cellules ductales ne durent que quelques semaines (Walker et al., 1992). Ces mêmes auteurs précisent bien que l'épithélium ductal serait constitué non pas de cellules acineuses différenciées, mais de cellules centroacineuses protodifférenciées.
Chez les lapins, au 15e jour post ligature notamment, des cellules claires dont le cytoplasme contient peu d'organites se trouvent intercalées dans l'épithélium pseudostratifié. Ces cellules claires pourraient correspondre soit à des cellules précurseurs soit à des cellules souches (oligopotentes) décrites par Rosenberg et al. (1988) chez le hamster diabétique au cours de processus de nésidioblastose et déjà visualisées en microscopie électronique chez les lapins P par Petkov et al. (1992).
Au niveau de la différenciation des cellules pancréatiques, l'existence et l'origine de la cellule souche canalaire est sujette à nombreuses controverses. Néanmoins, c'est après la pose de la ligature que nous avons pu constater que chaque cellule claire était située au pôle basal des ductules, contre la membrane constituée de collagène de type IV. En effet, lors de la prolifération, la potentialité d'être une cellule souche n'existe que pour les cellules qui sont en contact avec la membrane basale (comme cela a été démontré pour les cellules de l'épiderme et des cryptes intestinales). Cette position cellule/membrane basale est le facteur déterminant car la perte de contact déclenche la différenciation terminale (Alberts , 1990).
Chez le lapin, nous pouvons suggérer que lorsque ces cellules ont proliféré, elles vont se grouper en amas car il a déjà été rapporté au 15ème et au 30ème jour des amas de cellules endocrines à la périphérie des ductules après coloration au PAF (Petkov et al., 1992). Le fait que la membrane basale englobe à la fois le ductule et les cellules endocrines prolifératives, nous permet de dire que dans un amas les cellules proviennent d'un même ductule mais pas forcément d'une même cellule souche. Ces amas qui ont migré après séparation de l'épithélium ductal par poussée du tissu conjonctif, se constituent, au 90ème jour post ligature, en structures globulaires. En effet, ces cellules souches vont évoluer après mitose en cellules endocrines essentiellement A et surtout B (Petkov et al., 1992) qui ne sont pas encore biologiquement actives à ces périodes puisque nous n'avons pas pu les marquer par un anticorps anti-insuline monoclonal (Cortie et al., 1992).
Plus tardivement (180 jours), ces structures globulaires endocrines se regroupent pour former des clusters de différentes tailles dans lesquels persistent des cloisonnements constitués par des fibres de collagène. Ceci nous permet de dire que chez le lapin, se développent des processus de nésidioblastose sans toutefois pour l'instant préciser si cela aboutit à de véritables îlots, comme il est souvent signalé dans des pathologies semblables.
En effet, chez le hamster, après avoir engendré une fibrose pancréatique par enveloppement de la tête du pancréas avec un ruban de cellophane, Rosenberg et al. (1983) constatent également, à la deuxième semaine, l'induction d'une nésidioblastose, c'est-à-dire la formation de "nouveaux" îlots constitués de cellules A et B. Ce même phénomène de nésidioblastose a également été signalé chez la souris 5 jours après injection de streptozotocine. Les îlots qui apparaissent alors proviennent de la différenciation de l'épithélium ductulaire, deviennent hyperplasiques et envoient des interdigitations dans le parenchyme acineux adjacent (Papaccio et al., 1991).
Par ailleurs à long terme, chez les lapins ligaturés pendant 450 jours, nous avons pu visualiser en ultrastructure des cellules endocrines néoformées que nous avons appelées cellules mixtes, du fait de la présence dans leur cytoplasme de granulations a et ß côte-à-côte. Newman et al. (1986) ont eux aussi rapporté, chez des patients, que la nésidioblastose et l'adénomatose focale s'accompagnaient de la formation de cellules mixtes dites de type polycrine car elles étaient équipées des deux sortes de granules de sécrétion matures typiques a et ß. Ce même type de cellules apparaît également dans 77 % des adénocarcinomes pancréatiques ductulaires (Pour, 1988).
Par contre, chez les lapins ligaturés, nous n'avons jamais rencontré de cellules mixtes de type amphicrine, avec différenciation exocrine-endocrine, comme signalées dans des cas de nésidioblastose aussi bien que de nésidiodysplasie chez l'homme (Gould, 1983) ou que de carcinogénèse induite chez le hamster (Pour, 1989). Au niveau de ces cellules mixtes, on est obligé de s'interroger sur la présence "dans leur cytoplasme" de fibres de collagène organisées en faisceaux, disposés aussi bien longitudinalement que transversalement, fait dont on ne connaît pas de précédent. Ainsi, nous restons très réservés quant à une interprétation biologique par rapport à une possibilité de souffrance des tissus, impliquant une intrication cellules-fibres, et ceci selon un plan de coupe de cet enchevêtrement bien particulier. Cependant, nous suggérons une autre hypothèse qui repose sur une observation détaillée des images ultrastructurales. En effet, deux points ont retenu particulièrement notre attention:
Ces interruptions ne pourraient-elles pas
être soit la cause soit la conséquence de la
pénétration des fibres de collagène dans ces
cellules ? Un même processus a déjà
été décrit concernant l'infiltration de
microfibrilles amyloïdes entraînant la rupture de la
membrane plasmique de cellules B (Westermark et al., 1978). Rien ne
nous interdit de suggérer que, dans ces conditions, des
échanges de granulations a-ß puissent également
se produire consécutivement à ce
phénomène."
L'apoptose brève mais intense
des cellules acineuses pancréatiques est donc suivie de
phénomènes de proliférations et de
différenciations pouvant aboutir à la formation par
nésidioblastose de nouvelles cellules endocrines.
Apoptose des cellules endocrines pancréatiques
L'apoptose des cellules endocrines pancréatiques peut s'observer dans des conditions physiologiques normales. Ainsi, chez le rat, après accouchement, la masse des cellules B diminue. Cette diminution est causée par l'apoptose des cellules B pendant 4 à 6 jours après la mise bas (Scaglia & al., 1995). Le gène TRM2 et sa protéine, la clusterine, sont impliqués dans cette apoptose des cellules B ainsi que le gène TGFb1 dont l'expression augmente 3 jours après l'accouchement (alors que le niveau de TRM2 reste constant).
L'apoptose des cellules B peut également être induite par des facteurs alimentaires: chez le rat GK (modèle animal de DNID), une nourriture composée à 30% de sucrose induit une apoptose des cellules B caractérisée par de nombreuses atteintes oxydatives de l'ADN de ces cellules (Koyama & al., 1998). Ceci est a relier au phénomène de glucotoxicité observé chez Psammomys obesus, un modèle de DNID: en présence de fortes concentrations en glucose, les cellules B de cet animal subissent (in vitro) une phase transitoire de multiplication puis meurent par apoptose (Donath & al., 1999). L'existence de deux souches de cet animal réagissant différemment à l'exposition au glucose suggére l'existence de facteurs génétiques à l'origine de cette différence de"glucosensibilité".
Dans des situations pathologiques, l'apoptose des cellules B est impliquées dans le diabète de type I (Polychronakos & al., 1995). Dans ce cas, l'atteinte des cellules est due à une vaste famille de molécules (cytokines, acide nitrique, Streptozotocine, oxydants...) mais peut également être liée, chez des modéles animaux, à des facteurs alimentaires, en particulier à une alimentation fortement enrichie en protéines (Linn & al., 1999).
L'apoptose des cellules B peut également se révéler importante dans le diabète NID. En effet, les fibres amyloïdes qui s'accumulent tardivement dans le pancréas sont capables, au contact de leur membrane cellulaire, de déclencher l'apoptose des cellules B, contribuant ainsi à l'aggravation de l'état diabétique (Lorenzo & al., 1994). D'autres molécules sont capables de déclencher l'apoptose des cellules B, par exemple le TNF a (Stephens & al., 1999).
Il semble donc bien que le nombre de cellules B pancréatiques actives résulte en permanence d'un équilibre dynamique entre des mécanismes de replication (augmentant le nombre de cellules B) et d'apoptose (diminuant ce nombre). Cet équilibre a d'ailleurs été mis en évidence chez le rat par Bernard & al. (1999)
Toute perturbation de cet équilibre dynamique tendrait donc, sinon à déclencher, du moins à aggraver l'état diabétique.
De plus, le diabète de type II induit des phénomènes apoptotiques dans des organes éloignés du pancréas, par exemple le cerveau. Des neuropathies se retrouvent souvent chez des diabétiques de type II, et l'on doit remarquer qu'il existe dans le sérum de ces personnes un auto anticorps capable d'induire une apoptose des neurones (Srinivasan & al., 1998). Cette apoptose est calcium dépendante (elle est précédée d'une entrée massive de calcium dans la cellule). Cela est intéressant car une molécule déclenchant l'apoptose des cellules B pancréatiques, l'IAPP, est connu pour posséder justement des effets sur le métabolisme du calcium !
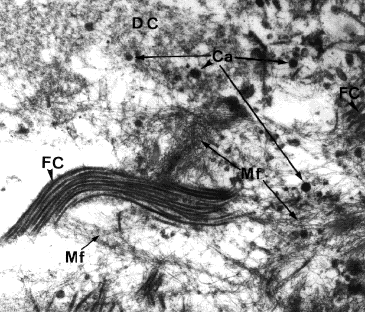
|
Cette molécule produite en abondance pendant le diabète NID possède un effet hypocalcémiant, et une accumulation de calcium a été détectée dans les cellules pancréatiques chez le lapin après 450 jours de ligature du canal pancréatique, voisinant avec des microfibrilles amyloïdes d'IAPP. ci contre: amas de calcium (Ca) voisinant avec des microfibres (Mf) amyloïdes et du collagène (FC) dans le tissu pancréatique d'un lapin après 450 jours de ligature du canal pancréatique. Photo C. Cortie |
 |
Rôle de l'apoptose dans le diabète
Une apoptose des cellules B provoque une insulinopénie. Or, ce phénomène est observé dans le diabète de type II. L'apoptose des cellules B peut être induite, nous l'avons vu, par des dépôts amyloïdes, mais pourrait également découler d'autres facteurs.
Ainsi, des produits de l'expression du gène STM2, impliqué dans une forme familiale de maladie d'Alzheimer, ont été identifiés dans le pancréas, le muscle squelettique et le coeur (Levy Lahad & al., 1996). L'expression de ce gène est particulièrement élevée dans le pancréas et le muscle (et non dans le cerveau). Ceci est surprenant car seul le cerveau est le siège de processus dégénératifs apoptotiques dans la maladie d' Alzheimer. Néanmoins, dans ces deux cas, une accumulation de fibres extracellulaires est impliquée dans des processus dégénératifs, de façon encore imparfaitement connue.
L'apoptose des cellules B observée dans le diabète de type I peut être prévenue par les molécules IGF (Polychronakos & al., 1995) et par la protéine antagoniste au récepteur de l'interleukine 1 (IL-1Ra - Giannoukakis & al., 1999). Il faudra sans doute à l'avenir rechercher dans quelle mesure les facteurs de croissance pourraient restaurer une population de cellules B fonctionnelles, en s'opposant à leur autodestruction. De même, des molécules comme la glutamine ou des antioxydant comme l'acide 1-pyrrolidinecarbodithioique (Caraher & al., 1999) ou la thioredoxine (Hotta & al., 1998) ont montré des effets antiapoptotiques dans ce type de diabète, on peut supposer qu'elles aurait une action similaire sur l'apoptose des cellules B du DNID.
Apoptose et complications du diabète
Ce sont les complications associées au diabète qui font toute la gravité de cette maladie. Les atteintes les plus morbides touchent les vaisseaux sanguins et le système nerveux central. A ces deux niveaux, on constate une exagération des phénomènes apoptotiques induits de façon indirecte par l'état diabétique.
Ainsi, une des complications induite par le diabète NID est la formation de plaques d'athéromes débouchant le plus souvent sur des cardiopathies obstructives aiguës. Il a été démontré (Fukumoto & al., 1998) qu'a ce niveau aussi des phénomènes apoptotiques sont impliqués.
Un Stress oxydatif meurtrier
L'hyperglycémie caractérisant
l'état diabétique et le stress oxydatif qu'elle
génère est également connu pour pouvoir
provoquer l'apoptose de neurones au niveau cérébral et
périphérique (Greene & al., 1999), ainsi que des atteintes variées sur les
processus de croissance de ces cellules et des structures qui y sont
associées (Russell & al., 1999). L'Insulin Growth Factor 1 possède
d'ailleur un effet protecteur sur ces neurones.L'effet du stress
oxydatif sur l'induction de l'apoptose des neurones a
été caractérisé in vivo et in vitro
(Russell, Sullivan & al.,
1999) et peut être reliés
à une activation de la caspase 3 par le glucose,
précédée par une altération des
crêtes mitochondriales.
L'apoptose étant sous la dépendance de signaux
intercellulaires comme les interleukines par exemple, il faudra mieux
appréhender les relations entre les différentes
cellules des îlots de Langherans, afin de mieux identifier les
signaux moléculaires qui maintiennent en vie les cellules B.
Plus que jamais, la vie d'un organisme pluricellulaire nous
apparaît comme un état transitoire résultant d'un
équilibre entre la pérennité des fonctions
cellulaires et le nécessaire contrôle de leur croissance
et de leur reproduction.